
S. Sikolia
Department of Botany, Maseno University, Kenya
J.C. Onyango
Department of Botany, Maseno University, Kenya
Research Journal of Botany
Year: 2009 | Volume: 4 | Issue: 1 | Page No.: 29-39







ABSTRACT
This study aims to elucidate the ontogeny and organization of the female gametophyte. Further, provide evidences for the strike phenomenon in the Podostemaceae. The female gametophyte ontogeny in Inversodicraea bifurcata Engl. I. keniensis sp. nov. Nagendran et Sikolia conforms to the Apinagia type of the monosporic category. The primary chalazal nucleus degenerates at the two-nucleate stage. The strike phenomenon met in the family is discussed. The organized female gametophyte is four-nucleate, four-celled and consists of two pear shaped synergids, a large central egg cell and a polar cell. Filiform apparatus are present in the synergids. Based on the female gametophyte ontogeny, there are no antipodal cell(s). Earlier reports of double fertilization are not confirmed, but only single fertilization take place in the family. The nucellar plasmodium is rationalized on its organization and ontogeny and is formed before fertilization in Inversodicraea. Effects of tension force and lytic enzymes during nucellar plasmodium organization and its ultimate significance are discussed.
PDF Abstract XML References Citation
How to cite this article
S. Sikolia and J.C. Onyango, 2009. Female Gametophyte in Two Kenyan Species of Inversodicraea
(Podostemaceae). Research Journal of Botany, 4: 29-39.
URL: https://scialert.net/abstract/?doi=rjb.2009.29.39
URL: https://scialert.net/abstract/?doi=rjb.2009.29.39
INTRODUCTION
Podostemaceae has been subdivided into Tristichoideae, Podostemoideae and Weddellinoideae subfamilies. Most studies of the Podostemaceae have focused attention on the shoot–root axis and female gametophyte development. The shoot and root axes development show the most enigmatic organs in Podostemaceae, usually deviating from the norm shoot apical and root apical meristems development pattern of the angiosperms (Koi and Kato, 2007). For example, there is no obvious shoot apical meristem in some species of Podostemoideae (Rutishauser, 1995, 2000; Rutishauser and Grubert, 1999), but occur in the Tristichoideae and Weddellinoideae (Koi et al., 2006). Similarly, there is variation in the root apical meristem and cap organization in Podostemaceae where the least specialized organization occurs in the Tristichoideae and Weddellinoideae (Koi et al., 2006).
The literature on the female gametophyte was reviewed by Battaglia (1971) and pointed out the need for reinvestigation in several taxa. Arekal and Nagendran (1975a, b, 1976, 1977a, b), Nagendran and Arekal (1976), Nagendran et al. (1977, 1980) studied the female gametophyte in many Indian taxa. Battaglia (1987) critically analyzed the observations and interpretations on the female gametophyte ontogeny. Battaglia (1987) categorically stated that all embryological data not assigned to the Apinagia type are doubtful cases and therefore require reinvestigation. Out of 73 species in 16 genera recorded for the African continent (approx. 26% of the total taxa known), only seven species representing three genera, Inversodicraea, Polypleurum and Tristicha have been studied so far. On the account of the endemism, inaccessibility for collection and availability of flowers and fruits only during a limited span of time, several taxa have not been investigated. Cusset and Cusset (1988) based on the morphological, anatomical, embryological and physiological data proposed a new class Podostenopsida beside the Magnoliopsida and the Liliopsida. In view of the above, it was decided to investigate the female gametophyte in two species of Podostemaceae available in Kenya.
MATERIALS AND METHODS
The study was carried out in the year 2000-2004. The plant material collected was fixed in Formalin-Acetic-Alcohol (FAA) and subsequently preserved in 70% ethanol. Fixed plant materials were taxonomically identified under a binocular dissecting stereomicroscope. The species are noted against their site of collection, below:
| • | Inversodicraea bifurcata Engler, along Chania river, 13-16 km upstream from Thika Town, Murang`a district, Central Province, Kenya |
| • | Inversodicraea keniensis sp .nov. Nagendran et Sikolia, along Chania river, 16 km upstream from Thika Town, Murang`a district, Central Province, Kenya, in the year 2000 |
| • | Two more trips, each at different period, an interval of week to the site were made to collect different developmental stages, of the specimen |
| • | For embryological study, individual flower buds of all sizes were isolated for each species (under a binocular dissecting stereomicroscope). They were dehydrated in the ethanol-xylol series and embedded in pure paraffin wax of 52°C melting point. Transverse and longitudinal sections were prepared at 7-12 μm m thick. Staining of sections in Heidehain`s Iron Alum-Hematoxylin using erythrosin or fast green in clove oil as counter stain, was done. DPX mountant was used to prepare permanent slides. Camera Lucida drawings of the female gametophyte stages were drawn at table level using a `Leitz` monocular microscope |
RESULTS
Inversodicraea Bifurcata
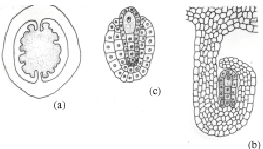
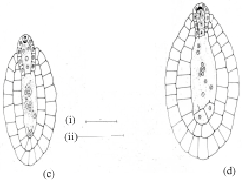
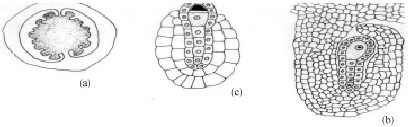
The gynoecium is bicarpellary and syncarpous. A superior, bilocular ovary bears numerous tenuinucellate, anatropous and bitegmic ovules on a swollen axile placenta in each locule. In a transverse section of a young ovary, several ovular primordial arise from the placental lump (Fig. 1a). Later, inside the developing ovule, a densely cytoplasmic large nucleated hypodermal archesporial cell differentiates (Fig. 1b). This cell without further division enlarges and transforms into the megaspore mother cell (Fig. 1c). It undergoes meiosis-I (Fig. 1d.). Of the resultant two dyad cells, the micropylar one degenerates regularly and is recognizable as a crescent-shaped cap at the micropylar end of the developing embryo sac (Fig. 1e).
The chalazal dyad cell undergoes the second meiotic division (Fig. 1f.). The two resulting nuclei move apart towards the poles of the embryo sac. A prominent vacuole is observed between these two nuclei (Fig. 1g). The chalazal megaspore nucleus begins to degenerates (Fig. 1h). Meanwhile, the micropylar megaspore nucleus enlarges in size (Fig. 1h). It undergoes the first mitotic division which is free nuclear (Fig. 1i). This results in a secondary two-nucleate embryo sac stage (Fig. 2a). The two nuclei undergo another free nuclear division (Fig. 2b). This results in a four-nucleate embryo sac (Fig. 2c). Wall formation results in organization of a four-celled embryo sac (Fig. 2d). The embryo sac consists of two pear shaped synergids, a large central egg cell and polar cell. Each of the synergids [was observed to] possess filiform apparatus (Fig. 2e). The development of the female gametophyte conforms to the Apinagia type, form B of Battaglia (1971).
| Fig. 1: | Early female gametophyte of Inversodicraea bifurcata. Bar I for a = 450 µm; Bar II for b-c = 50 µm |
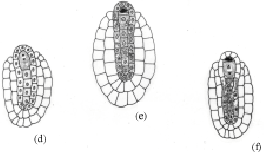
| Fig. 2: | Late female gametophyte of Inversodicraea bifurcata. Bar I for a, b, c and e Bar II for d = 40 µm) |
The pollen tube enters the ovule through the micropyle and crushes one of the synergids (Fig. 2e). The zygote formed after syngamy enlarges in size considerably (Fig. 2d). The degenerated polar cell and remnants of the pollen tube can be observed at this stage. The second male gamete is never discharged but remains in the cytoplasmic mass at the tip of the pollen tube (Fig. 2e). No fusion of the second male gamete and a polar cell was seen, implying that there is only single fertilization.
The nucellar situated below the megaspore mother cell in the developing embryo sac become densely protoplasmic (Fig. 1d-e) and elongate. When the female gametophyte is at the primary two-nucleate embryo sac stage, their walls breakdown and disappear (Fig. 1g). This phenomenon of wall disintegration commence at the chalazal end and proceeds micropylar-chalazal axis. The naked protoplasts start coalescing in the long cavity formed. Their nuclei often gather at different loci, in numbers less than eight, in the vacuolated cytoplasmic mass in the cavity. This transforms a structure consisting of multinucleate protoplasts in a long cavity and is referred to as the pseudo-embryo sac, observed below the embryo sac. It is organized before fertilization.
Inversodicraea keniensis
The gynoecium is bicarpellary and syncarpous. A superior bilocular ovary bears numerous tenuinucellate, anatropous and bitegmic ovules on axile placenta.
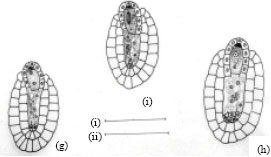
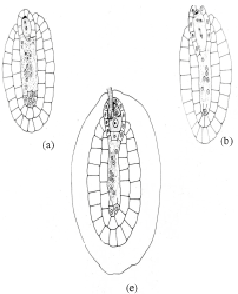
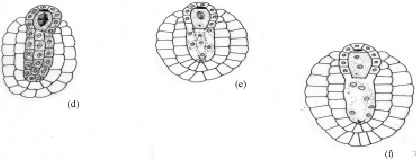
The ovular primordial develops from a placenta as protuberances (Fig. 3a). In the developing ovular primordium, a densely protoplasmic hypodermal archesporial cell differentiates early and transforms directly into the megaspore mother cell (Fig. 3b). The micropyle is organized by the outer integument only, as the inner one lags behind the former. After meiosis- I, the megaspore mother cell produces two dyad cells. The micropylar dyad cell degenerates and appears as an inverted cap of dark mass (Fig. 3c.).
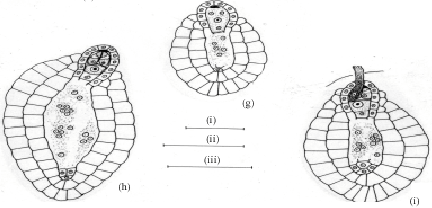
The nucleus of the chalazal dyad cell undergoes second meiotic division (Fig. 3d). Of the two megaspore nuclei produced, the chalazal one begins degeneration without further division (Fig. 3e). However, the other micropylar megaspore nucleus undergoes the first free nuclear mitotic division. Here, the degenerating chalazal megaspore nucleus seen as a picnotic structure disappears in subsequent stages (Fig. 3g-i). At late stages, the two micropylar megaspore nuclei have completed the second free nuclear mitotic division (Fig. 3g). Cellular organization in the four-nucleate embryo sac establishes two juxtaposed synergids, a large central egg cell and a polar cell (Fig. 3h). Filiform apparatus have been clearly observed in the synergids. The development of the female gametophyte conforms to the Apinagia type form B of Battaglia (1971).
After entering the ovule through the micropyle, the pollen tube penetrates the embryo sac destroying one of the synergids (Fig. 3i). The zygote formed after a male gamete fertilized the nucleus of the egg cell increases in size. The second male gamete is never discharged but can be seen retained at the tip of the pollen tube (Fig. 3i). Meanwhile, the polar cell below the zygote continues to reduce in size. At this stage, the disintegrated protoplasmic mass in the pollen tube overlapping the destroyed synergid components can be observed (Fig. 3i).
During the period of the developing embryo sac, the nucellar cells located below the female gametophyte become densely cytoplasmic and elongate (Fig. 3c-d). These cells are bound by a two-layered inner integument, that also, cover two third of the embryo sac in a chalazal-micropylar axis (Fig. 3c-i). The nuclei of the nucellar cells are large and distinct. Their walls remain intact before the primary two-nucleate embryo sac stage results (Fig. 3d). Later, their walls begin to breakdown (Fig. 3e) and disorganize. This process of wall disintegration begins at the chalazal end of the embryo sac and proceeds in a basipetal direction. The resultant-free protoplasts fuse one by one in the long cavity, thus formed. This happens before the secondary two-nucleate embryo sac stage (Fig. 3f). The number of protoplasts gathered at anyone loci in longisection are usually less than eight in the cavity. This, long cavity filled with multinucleate protoplasts, organize before fertilization (Fig. 3h). The entire structure formed below the embryo sac is the one usually referred to as the pseudo-embryo sac.
| Fig. 3: | Female gametophyte of Inversodicraea keniensis Bar I for a = 250 μm; Bar II for b-f and h = 45 μm, Bar III for g = 100 μm |
DISCUSSION
The gynoecium is syncarpous and bicarpellary in Inversodicraea bifurcata and I. Keniensis. A superior ovary bearing numerous minute ovules on a massive axile placenta is bilocular in Podostemoideae (Arekal and Nagendran, 1977b) of the Podostemaceae. Therefore, the reports of the ovary being unilocular with free central placentation (Rendle, 1925; Ramamurthy and Joseph, 1964) are not confirmed and are erroneous records for the family.
The ovular primordial arises on the placental hump as protuberances. In the longisection of an ovular primordium, four cells in axial row are seen in the nucellus. Of these, a hypodermal cell differentiates early in the ovule which bends in the form of a hook, causing its distal region to move closer to the placenta. Differentiation of the two integuments is simultaneous; but the outer one grows faster than the inner which lags behind. Thus, only the outer integument organizes the micropyle.
The differentiation of anatropous, bitegmic and tenuinucellate ovules from ovular primordial is almost uniform in the family. Later, well-defined and related patterns in the development and organization of the female gametophyte in different taxa of Podostemaceae follow. Conventionally, they are classified as the Apinagia type, the Dicraea type, the Podostemum type (Battaglia, 1971) and the Willisia type (Arekal and Nagendran, 1976). Each of these ontogenies commences as a densely cytoplasmic large nucleated hypodermal cell which differentiates early in the nucellus. It enlarges and directly transforms into a megaspore mother cell and remains at this stage until the level of water subsides in the streams in which these plants grow (Nagendran et al., 1977), exposing mostly the reproductive parts.
The megaspore mother cell undergoes asymmetric first meiotic divisions. The resultant dyad cells differ in size. The smaller micropylar one degenerates, without further division and is usually seen as an inverted cap of dark mass at the top of developing female gametophyte. It has no functional participation in the subsequent development and organization of the female gametophyte. On the contrary, the chalazal dyad cell is distinctively large with a prominent dense protoplasmic mass, functional in the second meiotic division of the ontogeny. The phenomenon of unequal first meiotic division is reported in other investigated taxa of Podostemaceae (Arekal and Nagendran, 1975a, b, 1976, 1977a, b; Nagendran and Arekal, 1976; Nagendran et al., 1976, 1977, 1980), Epipogium roseum (Arekal and Karanth, 1981).
The occasional atypical division (Meiosis-II) of the micropylar dyad cell (Magnus, 1913; Razi, 1949, 1955; Arekal and Nagendran, 1977a) or its persistence without further division (Jager-Zurn, 1967) is not confirmed but usually seen as an inverted cap of dark mass at the top of developing female gametophyte. It has no functional participation in the subsequent development and organization of the female gametophyte. This could be part of the abnormal cellular variations of the degenerating dyad cell phase. On the contrary, the chalazal dyad cell is distinctively large with a prominent dense protoplasmic mass, functional in the second meiotic division of the ontogeny. It is an indication of evolutionary process in the ontogenies of the Podostemaceae.
Arekal and Nagendran (1977a) observed that the nucleus of the non-functional dyad cell in the Zeylanidium olivaceum, may divide which may or may not be followed by a wall formation. Further, the second meiotic division of the chalazal dyad cell maintains its distinction by undergoing early division slightly before the micropylar one fully degenerates. The later dyad cell may be possessing callose deposited in its cell wall, because pollen tube enters on the side of the organizing female gametophyte. In view of the significance of unequal division in the process of differentiation, establishment of polarity becomes important factor during the division of the megaspore mother cell in determining the survival of the chalazal megaspore (or dyad cell). The nucellar wall disintegrates and could disrupt the course of nutrient flow and supply to the chalazal region of the developing megagametophyte. The wall disintegration commences during this primary nucleate stage.
Non-functional participation and inconsistency of the micropylar dyad cell during the developing megagametophyte fails to conform to the normal ontogeny of monosporic type because,…in the bisporic types, the upper dyad cell when dividing (Meiosis-II) give rise to a binucleate stage instead of the mononucleate cells as in monosporic type, (Battaglia, 1971). This concurs with the view that by the end of the first meiotic prophase the callose in the wall is usually thin or absent at the chalazal pole of the megaspore mother cell, in those species in which the germinal spore occurs at the chalazal positions whereas the micropylar pole of the megaspore mother cell has less callose in those species in which the micropylar spore forms the female gametophyte (Rodkiewicz and Bednara, 1976). And that, subsequent position of the germinal spore generally relates to the pattern of the callose deposition in the cell walls surrounding the megaspore nucleus in the developing female gametophyte (Haig, 1986). Given, callose deposition reduces the permeability of the nutrients and other essential metabolites to sustain the developing megagametophyte, then, the upper dyad cell is limited in further development. This type of ontogeny is seen in Oenothera sp. (Haig, 1986) where a correlation does exist between callose deposition and the germinal spore formed. Of significance, the situation in monosporic development shows tendency for the somatic spores or their derivatives being eliminated as non-functional during the ontogeny. Concurrently, the present investigations confirm that the micropylar dyad cell degenerates completely without subsequent morphological, embryological and functional significances in the megagametophyte ontogeny. However, the phenomenon of cell degeneration raises another question, does the chalazal dyad cell eliminate the micropylar one through competition within the developing megagametophyte? an observation reported in Oenothera biennis and O. suaveoleus (Noher de halac and Harte, 1975; Noher de halac, 1980). Further embryological investigations need to ascertain this contention.
The nucleus of the chalazal dyad cell completes the second meiotic division to produce two megaspore nuclei without cell wall formation between them. These nuclei are separated towards opposite poles of the female gametophyte by a vacuole. The report of cell wall formation at this two-nucleate megagametophyte stage in Polyplerum elongatum (Magnus, 1913) and Griffithella hookeriana (Razi, 1949) are not confirmed in the present study. This conforms to the observations for P. stylosum (Mukkada, 1964), P. dichotomum, P. filifolium and P. munnarensis (Nagendran et al., 1977). Until this primary two-nucleate phase, the female gametophyte development is specific in nature and fairly uniform in the family. This stage of megasporogenesis can be considered to be characterized with the normal number of meiotic divisions (two) but reduced and censored sporophytic genetic material. The other genetic component continues to disintegrate without functional role in the subsequent gametophyte ontogeny. The latter process means less utilization and considerable preservation of the maternal food resources. This is because of the reduced quantity and greater quality of the meiotic products in the course of the megasporophytic phase. Thus, megagametophytic phase ensue a censored genetic material (functional) and enough energy supply (preserved maternal metabolic substances-food resource) from the previous two stages of the ontogeny. This in itself shows an evolutionary process, exemplified by a high degree of reductionalism during the megasporophytic phase. This contention is an established fact in the various investigations because highly advanced megagametophytic ontogenies over a spell of time have evolved as progenies of the primitive-Polygonum type in diverse and advanced families of the angiosperm.
The ontogenies must be the result of several factors: polarity, course of nutrient supply and other aquatic factors, that come into interplay during a given ontogeny. Concurrently, either the megasporophytic or megagametophytic or in both phases, part of the whole or even the whole corresponding genetic material is made to be functional. This is natural mechanism for the existence and survival a given gametophyte ontogeny during adverse conditions and habitats, among the competing taxa. Thus, the theory of megaspore conflict of Haig (1986) suggests that the somatic spores or their derivatives having a role in successful gametophyte function have a tendency to become non-functional. This may ascertain the strike phenomenon of Davis (1966), reported in Podostemaceae (Nagendran et al., 1977, 1980) and confirmed to be taking place in the post-meiotic mitosis in the present study. This is exhibited by the total disintegration of the primary chalazal nucleus which commences after the cell wall of the nucellar cells disorganize, below the female gametophyte. This nucleus completely disappears as soon as a distinct vacuole separating it from the micropylar megaspore one becomes invisible. Similar phenomenon does occur soon prior to the cellularization of the megagametophyte starts for the investigated taxa of Tristichoideae (Arekal and Nagendran, 1976). This strike phenomenon has been reported in Epipogium roseum (Arekal and Karanth, 1981); but it is partially or complete, in similar biological specialized family, Orchidaceae (Abe, 1972).
The primary chalazal nucleus is positioned in the upper intersection region of the inner integuments (i.e., which depicts no nutritional value) but below the attachment of the outer integuments to the female gametophyte. On the contrary, the micropylar megaspore nucleus position itself in the total interspaced area of the upper outer integuments, just below the fully degenerated upper dyad cell which may have a supplementary food resource region (Wilms, 1980). The implication is that the micropylar megaspore nucleus is well-placed with respect to the food supply source distant from the possible lytic action of enzymes of the crushed nucellar cells, compared to the primary chalazal nucleus. Concomitantly, Battaglia (1971, 1987) pointed out that the chalazal region may be physiological depressed or hypofunctional. This can be explained because the chalazal megaspore nucleus becomes disadvantaged in placement towards the direction and source of the nutrient supply unlike the micropylar one which is in closer proximity with the outer integuments that contain starch. Therefore, the simple metabolite substances easily diffuse into its interspaced area.
Evidently, the primary chalazal nucleus degeneration is compatible with its reduction in size vis-� -vis its disappearance. This is an indication of the possible reduction in physiological activities (Battaglia, 1971). This may occur as a result of the change in the course of nutrient supply, as observed in Spinacia sp. (Wilms, 1980) where a similar course of ontogeny depicts the degeneration of the primary chalazal nucleus. This is in contrast to the morphological orientation of the primary micropylar nucleus, which is in the central area of the megagametophyte and receiving substantial food supply. Therefore, the chalazal megaspore nucleus division must be a quite uncommon event. However, the functional state of the micropylar megaspore nucleus is supportive because it does maintain its distinctive embryological image by undergoing division slightly before the degeneration of the chalazal one sets-in. Then, how can the antipodal cell(s) be organized if its precursor- the primary chalazal nucleus completely disintegrates without further division during the ontogeny? Accordingly, only the primary micropylar nucleus, through two free nuclear divisions, contributes to all the four nuclei in the organized female gametophyte. Consequently, the micropylar quartet of nuclei can only organize into two synergids, an egg cell and a polar cell. The latter cell considerably diminishes in size to a small dark mass, which can be an equivalent of its remnant nucleus. Each of the synergids possesses filiform apparatus. This clearly ascertains the morphological orientation of the female gametophyte. The present investigations confirm the reports of Nagendran et al. (1980) regarding complete absence of the antipodal cell in the female gametophyte consisting of two synergids, a large central cell and a polar cell; contrary to the view of Kapil and Bhatnagar (1978). Thus, the concept that, ……. Since the days of Hofmeister and Strasburger …. The antipodal cell (Strasburger`s Antipoden oder Genenfusslerinen) is a cell situated at the chalazal end of the mature ES, usually 1-nucleate, regularly degenerating at fertilization or, rarely, showing a mitotic activity (phenomenon of polyantipody), (Battaglia, 1987), could be admissible only if it is a derivative of the primary chalazal nucleus during the female gametophyte ontogeny. It follows that the status of any component in the organized female gametophyte must reflect its ontogenetic origin at megagametogenesis and its functional role before its morphological location is considered to be requisite as a matter of functional activity, fact and precision. This is the case, even if reverse polarity or other factors plays a role in determining the course of the ontogeny in other taxa of the angiosperm. Therefore, the development and organization in the present taxa of Inversodicraea conform to the Apinagia type, form B of Battaglia (1971).
Nagendran (1974) reinterpreted Apinagia type as monosporic and not reduced bisporic embryo sac development as earlier embryologists who investigated this type of female gametophyte considered. This classification (Nagendran, 1974) corroborates the present observations that only the micropylar megaspore nucleus contributes all the four nuclei which alone participate in the cellular organization of the mature female gametophyte. This concurs with the criterion explicitly expressed in the original definition of classifying female gametophyte in the angiosperms by Maheshwari (1937) and pointed out by Nagendran (1974) that …… an embryo sac formed from the divisions of a single megaspore nucleus should be called monosporic; and when the two take part in its development, it is bisporic; and when all the four contribute to it, it is tetrasporic p.306. Due to the strike phenomenon the chalazal megaspore nucleus is eliminated in the ontogeny. This is also reported in other investigated taxa of Podostemaceae (Mukkada, 1969; Arekal and Nagendran, 1977b; Nagendran et al., 1977). It is prudent that, the number of megaspore nuclei at the prime stage of the megagametophyte formation, its nuclear mitotic products and their subsequent participation in the cellular organization of mature female gametophyte needs to be considered while classifying its types in the angiosperms. Then, the Apinagia type is justified to be reinterpreted as monosporic (Nagendran, 1974) because only one megaspore nucleus undergoes the first and second nuclear mitotic divisions producing all the four nuclei that alone participate in the cellularization and are present in an organized female gametophyte.
The contention of Battaglia (1987) that all the embryological cases not assigned to the Apinagia type are …….. Doubtful cases …….. appear to be drastic and may not be true, because many embryologists after investigations and/or reinvestigations have independently confirmed their existence. The Podostemum type has been observed in Farmeria metzgeriodes (Trimen) Willis, Podostemum subulatus Gardn. (Magnus, 1913), Weddellina squamulosa Tul., (Chiarugi, 1933), Hydrobryopsis sessilis (Willis) Engler (Arekal and Nagendran, 1975b), the Polypleurum type in Polypleurum agharkarii (Nandi) (Nagendran et al., 1977), P. dichotomum (Gardn.) Hall (Nagendran et al., 1977), P. elongatum (Gardn.) Hall (Magnus, 1913; Mukkada, 1964; Nagendran et al., 1977), P. munnarensis Arekal et Nagendran (Nagendran et al., 1977), P. Stylosum (Night) Hall (Mukkada, 1964; Nagendran et al., 1977), the Willisia type in Willisia selaginoides (Bedd.) Warming ex Willis (Arekal and Nagendran, 1976). In all the female gametophyte types, Strike phenomenon has been recorded to be the consequence leading to primary chalazal nucleus vis-a-vis its products, elimination during the ontogeny in the family. The micropylar quartet of nuclei organizes and results in the corresponding, the Apinagia type, form B; and occasionally the Apinagia type, form A of Battaglia (1971). However, apart from the high degree of reduction compounded by specialization in the ontogenies of the Apinagia type, the Podostemum type (considered doubtful by Nagendran et al., 1977), the Polypleurum type and the Willisia type, the organization and morphological orientation of the Polypleurum type (Nagendran et al., 1977) is rationalized on the basis of reverse polarity. Further, Battaglia (1971, 1987) has not investigated any taxon to support his contention for the type of the female gametophyte development in the family. The work (questions and doubts) of Battaglia (1971, 1987) is only analytical. Too, of the four nuclei of the micropylar quartet of nuclei, polarity is well expressed when the central cell give rise to the central egg cell. From the present investigations, the ontogeny of the Apinagia type does exist; and since it seems to be of major occurrence from which the others are derived; and that the reduction process (strike phenomenon) and specialization phenomena does exist in it, the same can be attributed to the other ontogenies of the Podostemaceae. But lack of evidences using Transmission electron microscope in the investigations (Battaglia, 1987) in determining the fate of the primary chalazal nucleus and its products needs to be undertaken for further clarification.
Fertilization is porogamous. The pollen tube penetrates the female gametophyte destroying one of the synergid on its pathway, contrary to Razi`s (1955) observation between the two synergids. The other synergids only degenerate during embryogenic stages. The present mode of pollen tube pathway has been recorded in Terniola zeylanica (Gardn.) Tul., (Chopra and Mukkada, 1966), Indotristicha ramosissima (Wight) van Royen (Mukkada, 1969) and Mourera fluviatilis Aublet (Went, 1908). The contents of the pollen tube are overlapped with the protoplasmic mass of the destroyed synergid and are often visible as a dark mass with two nuclei in a degenerated state. One of these nuclei may be the non-discharged second male gamete. These observations confirm the reports of earlier investigators (Went, 1908; Chopra and Mukkada, 1966; Mukkada, 1969), that the second male gamete is not discharged in the Podostemaceae family. According to Mukkada (1969), often the tip of the pollen tube and one of the synergids persist as late as quadrant and octant stages of the pre-embryo. At this time, the polar cell is degenerated or absent, a process which commences before in other taxa in the family (Arekal and Nagendran, 1975a ,1977a, 1977b; Nagendran et al., 1976). The chances of the polar cell being fertilized by the second male gamete do not arise because none of the microscopic preparations showed the fusion. Further, there is no endosperm in the family and the primary endosperm nucleus never forms in the investigated taxa of Podostemaceae (Mukkada, 1969; Arekal and Nagendran., 1977a; Nagendran et al., 1976). The present study contrasts the reported fusion of the second male gamete with a polar nucleus by Razi (1955). There is no double fertilization in the studied taxa. The function of the endosperm, in other families of angiosperms, is compensated by the nucellar plasmodium in the Podostemaceae. The egg cell fertilized by the first male gamete produces the zygote which increases in size considerably and one-layered cell wall with a distinct large nucleus (seen as dark, rounded, at the central of the cell) that is two-thirds of the zygotic stage in earlier phases; and extends towards the chalazal region of the female gametophyte as well as sideways. This confirms a similar type of fertilization recorded in the family (Went, 1908; Chopra and Mukkada, 1966; Mukkada, 1969; Arekal and Nagendran, 1975b, 1977a, b; Nagendran et al., 1976). Thus, only single-fertilization takes place in the family.
REFERENCES
- Arekal, G.D. and K.A. Karanth, 1981. The embryology of Epipogium roseum (Orchidaceae). Plant Syst. Evol., 138: 1-7.
CrossRef - Arekal, G.D. and C.R. Nagendran, 1975. Embryo sac of Hydrobryopsis sessilis (Podostemaceae) origin, organization and significance. Bot. Notiser, 128: 332-338.
PubMed - Arekal, G.D. and C. R. Nagendran, 1976. A new type of embryo sac organization in Angiosperms. Curr. Sci., 45: 717-719.
PubMed - Arekal, G.D. and C.R. Nagendran, 1977. Female gametophyte in Zeylanidium (Podostemaceae). A clarification. Phytomorphology, 27: 123-129.
PubMed - Battaglia, E., 1971. The embryo sac of podostemaceae. An interpretation. Caryologia, 24: 403-420.
PubMed - Chopra, R.N. and A.J. Mukkada, 1966. Gametogenesis and Pseudo-embryo sac in Indotristicha ramosissima (Wight) Van Royen. Phytomorphology, 16: 182-188.
PubMed - Cusset, G. and C. Cusset, 1988. Etude sur les Podostemales. 10. Structures florales et végétatives des Tristichaceae. Bulletin du Muséum National d'Histoire Naturelle Paris, 10: 179-218.
PubMed - Jager-Zurn, I., 1967. Embryologische untersuchungen and vier Podostemaceen. Osterr. Bot. Z., 114: 20-45.
CrossRef - Koi, S. and M. Kato, 2007. Developmental morphology of the shoot in Weddellina squamulosa and implications for shoot evolution in the Podostemaceae. Ann. Bot., 99: 1121-1130.
Direct Link - Koi, S., R. Fujinami, N. Kubo, R.I. Tsukamoto, R. Imaichi and M. Kato, 2006. Comparative anatomy of root meristem and root cap in some species of Podostemaceae and evolution of root dorsiventrality. Am. J. Bot., 93: 682-692.
Direct Link - Mukkada, A.J., 1969. Some aspects of the morphology, embryology and biology of Terniola zeylanica (Gardner) Tulasne. New Phytol., 68: 1145-1158.
CrossRef - Maheshwari, P., 1937. A critical review of the types of embryo sacs in angiosperms. New Phytol., 36: 359-417.
Direct Link - Nagendran, C.R. and Arekal, G.D., 1976. The embryo sac of Griffithella hookeriana. A re-investigation. Phytologist, 26: 359-363.
PubMed - Nagendran, C.R., V.V. Anand and G.D. Arekal, 1980. The embryo sac of Podostemum subulatus (Podostemaceae). A re-investigation. Plant Syst. Evol., 134: 121-125.
CrossRef - Nagendran, C.R., G.D. Arekal and K. Subramanyam, 1977. Embryo sac studies in three Indian species of Polypleurum (Podostemaceae). Plant Syst. Evol., 128: 215-226.
CrossRef - Nagendran, C.R., K. Subramanyam and G.D. Arekal, 1976. Development of the female gametophyte in Hydrobryum griffithii (Podostemaceae). Ann. Bot., 40: 511-513.
Direct Link - Noher de Halac, I., 1980. Fine structure of the nucellar cells during development of theembryo sac in Oenothera biennis L. Ann. Bot., 45: 515-521.
Direct Link - Noher de Halac, I. and C. Harte, 1975. Female Gametophyte Competence in Relation to Polarization Phenomenon During the Megagametogenesis and Development of the Embryo Sac in the Genus Oenothera. In: Gamete Competition in Plants and Animals, Mulcahy, D.L. (Ed.). North-Holland Publishers, Amsterdam, pp: 43-56.
- Rutishauser, R., 1995. Developmental patterns of leaves in Podostemaceae compared with more typical flowering plants: Saltational evolution and fuzzy morphology. Can. J. Bot., 73: 1305-1317.
CrossRef - Rutishauser, R. and M. Grubert, 1999. The architecture of Mourera fluviatilis (Podostemaceae): Developmental morphology of inflorescences, flowers and seedlings. Am. J. Bot., 86: 907-922.
PubMed - Razi, B.A., 1949. Embryological studies of two members of the Podostemaceae. Bot. Gaz., 111: 211-218.
PubMed - Rodkiewicz, B. and J. Bednara, 1976. Cell wall ingrowth and callose distribution in megasporogenesis in some Orchidaceae. Phytomorphology, 26: 2276-2281.
PubMed - Wilms, H.J., 1980. Development and composition of the Spinach ovule. Acta Bot. Neerl., 29: 243-260.
Direct Link