Hot Coffee: The Identity, Climate Profiles, Agronomy, and Beverage Characteristics of Coffea racemosa and C. zanguebariae
 Aaron P. Davis1*
Aaron P. Davis1*  Roberta Gargiulo1
Roberta Gargiulo1  Iolanda N. das M. Almeida2
Iolanda N. das M. Almeida2  Marcelino Inácio Caravela3
Marcelino Inácio Caravela3  Charles Denison4
Charles Denison4  Justin Moat1
Justin Moat1- 1Royal Botanic Gardens, Kew, Richmond, United Kingdom
- 2Cabo Delgado Provincial Directorate of Culture and Tourism, Pemba, Mozambique
- 3Faculty of Natural Science, University of Lúrio, Pemba, Mozambique
- 4Cultivar Coffee, Dolphin Coast, South Africa
Climate change poses a considerable challenge for coffee farming, due to increasing temperatures, worsening weather perturbations, and shifts in the quantity and timing of precipitation. Of the actions required for ensuring climate resilience for coffee, changing the crop itself is paramount, and this may have to include using alternative coffee crop species. In this study we use a multidisciplinary approach to elucidate the identity, distribution, and attributes, of two minor coffee crop species from East Africa: Coffea racemosa and C. zanguebariae. Using DNA sequencing and morphology, we elucidate their phylogenetic relationships and confirm that they represent two distinct but closely related species. Climate profiling is used to understand their basic climatic requirements, which are compared to those of Arabica (C. arabica) and robusta (C. canephora) coffee. Basic agronomic data (including yield) and sensory information are provided and evaluated. Coffea racemosa and C. zanguebariae possess useful traits for coffee crop plant development, particularly heat tolerance, low precipitation requirement, high precipitation seasonality (dry season tolerance) and rapid fruit development (c. 4 months flowering to mature fruit). These attributes would be best accessed via breeding programs, although these species also have niche-market potential, particularly after further pre-farm selection and post-harvest optimization.
Introduction
Coffee farming and the success of the coffee value chain is constrained by numerous challenges, including fluctuating market prices, adverse weather, pests and diseases, and various social challenges. Anthropogenic climate change has a compounding influence on these factors (IPCC, 2014) and has been considered as the single overarching challenge facing long-term sustainability for coffee production (Davis et al., 2019). There are three main adaptation strategies for ensuring resilience for the coffee farming sector under climate change (Davis et al., 2021): (1) farm adaptation, by changing the air and soil microclimate on the farm, via various means; (2) relocation of coffee farming areas, e.g., to higher, cooler elevations; and (3) replacement, i.e., changing the crop using new cultivars, new hybrids, or alternative species. All three strategies have merit, but relocation (2) and replacement (3) offer the most potential over the longer term, under existing climate change projections (Davis et al., 2021). Despite the substantial potential of relocation for countries with suitable elevational range, such as Ethiopia (Moat et al., 2017), or other forms of spatio-climatic intervention, there are likely to be issues around competing land-use (for communities and biodiversity) and other negative societal implications. There will no doubt be winners and losers as farms either diversify into coffee or move away from coffee, as the climate changes. The capacity for human migration is usually limited and almost certainly undesirable. At this time, changing the crop therefore represents the most suitable adaptation pathway for coffee under accelerated climate change (Davis et al., 2019, 2021). Our current coffee crop species, Arabica (Coffea arabica) and robusta (C. canephora) still provide ample coffee to supply the global value chain, but narrative from farmers across the world's coffee belt, and ongoing shortfalls in production during weather perturbations and cyclic climatic phenomena, tell of ever-increasing climate-related issues. This is a result of the specificity of the climate envelopes for Arabica (Moat et al., 2017, 2019; Davis et al., 2018, 2021) and robusta (Kath et al., 2020), the perennial nature of the crop, and the fact that coffee farming has been extended into suboptimal climatic space for these two species and their cultivars.
Amongst the 130 wild coffee species (Davis and Rakotonasolo, 2021) there could be potential amongst the underutilized species (e.g., C. liberica), minor crop species (e.g., C. eugenioides, C. racemosa) or the (numerous) non-utilized species, to broaden the coffee crop portfolio, and provide material for coffee crop development (Davis et al., 2019) in an era of accelerated climate change. There are 42 priority coffee species (Davis et al., 2019), worthy of further evaluation and experiment. The case of C. stenophylla, which has an Arabica-like taste but can grow successfully at much warmer temperatures (mean annual temperature requirement 6.2–6.8°C higher than Arabica; Davis et al., 2021), demonstrates the potential of other coffee species to prove climate resiliency crop alternatives or, perhaps more importantly, resources for breeding. Even if C. stenophylla coffee proves to be successful, there will still be a requirement to further broaden the climatic portfolio of coffee crop species, including those that can grow and crop under both warmer, drier and more seasonal conditions than Arabica and robusta, and even C. stenophylla. In this respect, the current study focused on two minor coffee crops of East Africa, viz. C. racemosa and C. zanguebariae.
Coffea racemosa was described by Loureiro (1790) based on a herbarium specimen collected in Mozambique, although this specimen has not been recovered and is now replaced by a new type (neotype) specimen from the Massingir District (Gaza Province) in southwestern Mozambique (Bridson, 2003). Given its rather unique appearance, and knowledge of its use as a crop from previous times, C. racemosa has been adequately covered in many classical works on coffee (e.g., Cheney, 1925; Wellman, 1961; Haarer, 1962) but is notably absent from the work of Cramer (1957) possibly because Java does not provide a favourable environment for the growth of C. racemosa. A thorough review of C. racemosa was provided by Guerreiro Filho (1992), covering botanical features, taxonomic classification, geography (distribution), genetics and breeding, chemistry, possible attributes for the improvement (pest and disease resistance, climate resilience, shortening of fruiting time compared to Arabica and robusta), and a brief overview of sensory qualities (flavour). Attributes of particular note reported by Guerreiro Filho (1992) were: tolerance of higher temperatures and low monthly precipitation values, mostly based on the findings of Krug (1965) and Halle and Faria (1973); hybridization success with Arabica and robusta; resistance to various coffee pests, particularly coffee leaf miner (Perileucoptera coffeella) and nematodes (Meloidogyne species) and disease; and reduction in fruiting times for interspecies hybrids. According to (Guerreiro Filho, 1992), broad scale success of C. racemosa as a crop species was deemed unlikely due to its very small seed (coffee bean) size, based on examination and data from Lopes (1974), which stated a mean seed size of c. 6.3 × 5.1 × 3.2 (length, width, thickness). No yield (production) figures were given by Guerreiro Filho (1992) but this is also, of course, a key factor for agronomic and value chain success.
Climate information provided for C. racemosa by Krug (1965) and illustrated by Guerreiro Filho (1992; Figure 1, areas a, b, c) reported: (a) mean monthly temperatures of 18.9–26.4°C, an annual precipitation of c. 1,600 mm, and a 4–5 month dry season (May to October), for populations occurring at 200–500 m in central Mozambique; (b) mean monthly temperatures of 21.7–28.3°C, and 1,200–1,300 mm annual precipitation for low elevation (0–200 m) coastal areas of central Mozambique; and 19.6–27.3°C, and a 700–900 mm annual precipitation, for populations in areas in low elevation (0–200 m) coastal areas of southern Mozambique. Halle and Faria (1973) reported that between latitude 22°S and 25°S, at an elevation not exceeding 40 m, the annual precipitation for C. racemosa was 647 mm. Overall, these climate profiling data are useful but somewhat contradictory for a single species, particularly given that the mean annual precipitation requirement for C. arabica and C. canephora is around 1,600 m (Davis et al., 2021).
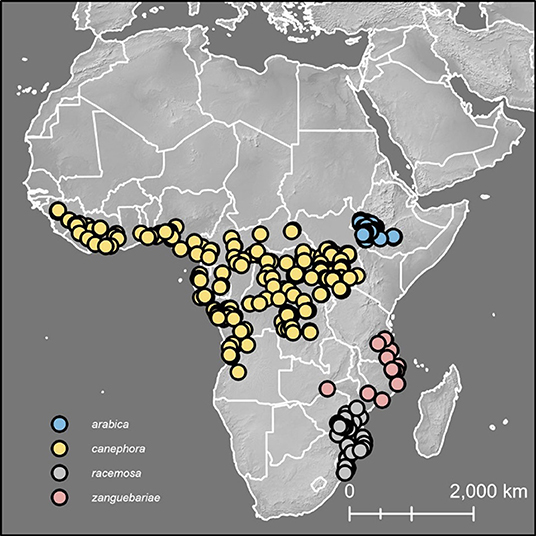
Figure 1. Distribution map for C. arabica, C. canephora, C. racemosa and C. zanguebariae. Figure also indicates ground point data for climate profiling analysis (Figures 8, 9).
Coffea zanguebariae was also described by Loureiro (1790), based on material collected from Zanzibar, which Loureiro remarks to have been taken there by the Portuguese. Due to similarities in morphology, C. zanguebariae has been routinely confused with C. racemosa (Haarer, 1962; Guerreiro Filho, 1992), other coffee species (Cheney, 1925; Haarer, 1962), and its identity has been undoubtedly mistaken (Cramer, 1957). Chevalier (1942, 1947) was well acquainted with C. racemosa but included what we now know as other Coffea species under C. zanguebariae, and most notably C. pseudozanguebariae (Bridson, 1982); this has led to considerable confusion over the identity of this species (Laíns E Silva, 1954; Guerreiro Filho, 1992). Indeed, Guerreiro Filho (1992) clearly considered C. racemosa and C. zanguebariae to be represented by a single species. On the basis of morpho-taxonomic study, Bridson (1988, 2003) reassessed the circumscriptions of C. zanguebariae and C. racemosa, demonstrated that they were separate species, and resolved outstanding taxonomic and nomenclatural issues. Botanists working in Mozambique have generally followed the assessment of Bridson (e.g., Burrows et al., 2018). Bridson (2003) considered C. racemosa as indigenous to central and southern Mozambique, northern South Africa (Kwa Zulu Natal), and eastern Zimbabwe, as a plant of coastal forest, riverine forest, deciduous woodland and bushland at elevation of 0–500 m. Coffea zanguebariae was reported by Bridson (1988, 2003) as indigenous to southern Tanzania, northern Zimbabwe and northern Mozambique, occurring in dry deciduous forest, and riverine and coastal thicket, at an elevation of 10–350 m. Common synonyms given by Bridson (2003) for C. racemosa include C. mozambiciana, C. ramosa, and C. swynnertonii; and for C. zanguebariae, synonyms include C. ibo, C. schumanniana, and C. zanzibarensis. These names become important when tracing the history of these species in cultivation and commerce.
Despite the apparent taxonomic and nomenclatural clarity afforded for C. racemosa and C. zanguebariae by Bridson (1988, 2003), there remains a great deal of confusion between these species in commerce and in coffee research. The phylogenetic placement of C. zanguebariae remains inconclusive (Maurin et al., 2007; Davis et al., 2011); and this species has not been included amongst recent molecular sampling for the genus (e.g., Hamon et al., 2017). With respect to the possible development of C. racemosa and C. zanguebariae as either coffee crops or plant breeding resources, certainty over the circumscription of these species, and understanding the relationship between them and other Coffea species, is required.
In addition to the incongruences reported for climate profiles and systematic uncertainty, there is a third point of contention regarding C. racemosa and C. zanguebariae, viz. their flavour characteristics. In the review by (Guerreiro Filho, 1992) various reports are given for the flavour quality of C. racemosa. According to Silva (1956) C. racemosa provides an aromatic drink, of slightly dark colour and low caffeine; Chevalier (1947) noted from a herbarium sheet than the “Grain is considered analogous to Moka [C. arabica], it is better than any other;” and Carvalho (pers. comm. in Guerreiro Filho, 1992) reported an inferior quality to that of C. arabica. Other reports include that of Haarer (1962) who stated that: “Coffee prepared from the beans [of C. racemosa] is used by the local settlers and said to be of excellent quality,” and Laíns E Silva (1954) that it produced an aromatic, lightly coloured beverage, with a low caffeine content, and was much favoured for batch mixing with C. arabica, but that this coffee was sought after in Mozambique without mixing. Published information, or otherwise, on the sensory qualities of C. zanguebariae appears to be non-existent.
Further points of ambiguity are apparent. Guerreiro Filho (1992; Figure 1) provide some basic details on the distribution of cultivated (farmed) C. racemosa (including C. zanguebariae; see above) but the distinction between cultivated and wild distributions was not made. The vernacular name “Inhambane coffee” is reported from several herbarium specimens of C. racemosa farmed in the region of that name in southern Mozambique (Bridson, 2003). There are early reports of the cultivation of C. zanguebariae: Cheney (1925) remarked that C. ibo (i.e., C. zanguebariae fide Bridson, 2003), appeared on the German southeast African market in 1893, and on Ibo island (north-eastern Mozambique) it was being grown in the 1920s as a substitute for C. arabica.
There remains, therefore, five main points of ambiguity for C. racemosa and C. zanguebariae, viz.: the systematic (phylogenetic) relationship between these two species, and other East Africa taxa; their climatic (and agro-climatic) requirements; their sensory qualities; their current usage as minor crops. An obvious shortfall for cultivated C. racemosa and C. zanguebariae is basic agronomic knowledge, such as harvest periods, yield, and conversion ratios to green coffee. Our objectives were, therefore, to: (1) elucidate the phylogenetic and spatio-phylogenetic relationship between C. racemosa and C. zanguebariae and to confirm or refute their existence as separate species; (2) model the climate profiles for these species, with a view to gaining a better understanding of their climatic requirements and agro-climatic suitability; (3) to provide preliminary sensory (flavour) information; and (4) to provide basic agronomic data. To undertake these objectives, we utilized plastid and ITS sequence data, climate profiling analyses using ground-point data, field and herbarium observation (of wild and farmed plants), farm surveys, literature review, and preliminary sensory information on beverage quality using standard industry procedures and narratives from coffee tasting professionals.
Methods
Fieldwork and Other Observation Data
Farm visits were made between 2016 and 2020, in northern South Africa (Kwa-Zulu Natal), at Hluhluwe (for C. racemosa) and in north-eastern Mozambique (Cabo Delgado Province), on Ibo Island and Quirimba Island and near Pemba on the mainland (for C. zanguebariae). Study of wild populations was undertaken north of Durban in South Africa (C. racemosa) and west of Pemba in north-eastern Mozambique (C. zanguebariae). Location, morphology, ecology, and agronomy data were also collected in situ. Herbarium specimens were consulted from five herbaria [BM, BR, K, P, WAG; codes follow Holmgren et al. (1990) and Thiers (2019)] for additional information, including habitat, vegetation, environment, morphology, cultivation history and trade.
Data for Mapping and Climate Profiling
Ground points derived from herbarium specimens and field survey were used to provide the data for the climate profiling analyses and to produce distribution maps for indigenous locations of C. arabica and C. canephora, and indigenous and cultivated (farmed) locations of C. racemosa and C. zanguebariae. Herbarium data are well suited for providing these data because they are verifiable in space (location), time (date) and form (species identity). We consulted herbarium specimen records from five herbaria (BM, BR, K, P, WAG). Location data from the herbarium specimens were georeferenced (if not already available), manually checked for geolocation accuracy (1 km or less) using GoogleEath® and corrected if necessary. For the distribution maps and climate profiling analysis we used a dataset of 1,087 ground point records, comprising 711 records for C. arabica, 304 for C. canephora, 50 for C. racemosa, and 22 for C. zanguebariae. We used indigenous and cultivated datapoints for C. racemosa and C. zanguebariae as they were either overlapped or contiguous. Disaggregated in this way, the number of datapoints for each class was 36 wild and 14 cultivated for C. racemosa, and 15 wild and seven cultivated for C. zanguebariae.
Distribution maps were produced in ArcGIS Pro 2.6.1 [Environmental Systems Research Institute (Esri), 2020], using background and country data from Natural Earth (https://www.naturalearthdata.com/).
Climate Profiling Analyses
For these analyses we resampled all specimen data to remove duplicates within 1 km of each other, reducing the total number of records used from 1,087 to 463 (193 for C. arabica, 200 for C. canephora, 50 for C. racemosa, and 20 for C. zanguebariae. To understand the fundamental climatic requirements, the statistics package R (R Core Team, 2016) was used to sample specimen data against 19 Bioclim variables (Busby, 1991) from the CHELSA dataset (Karger et al., 2017). For an overview of climatic parameters, we selected the following three Bioclims: Bio1 Annual mean temperature, Bio12 Annual Precipitation, and Bio15 Precipitation Seasonality. Scatter and density plots were plotted using R (R Core Team, 2016), using the ggplot2 (Wickham, 2016) and ggpubr packages (Kassambara, 2020). For validation purposes, our modelled annual temperatures (from Bio 1), annual precipitation (Bio12) and precipitation seasonality (Bio15) used to produce Figures 1, 2 were compared against publicly available monthly mean temperature precipitation charts for East Africa and published data (Laíns E Silva, 1954; Alègre, 1959; DaMatta, 2004; DaMatta and Ramalho, 2006; Kath et al., 2020). Our modelling methods have been shown to provide climate metrics that are similar to those provided for coffee species in cultivation, produced by direct measurement and other means (Davis et al., 2021).
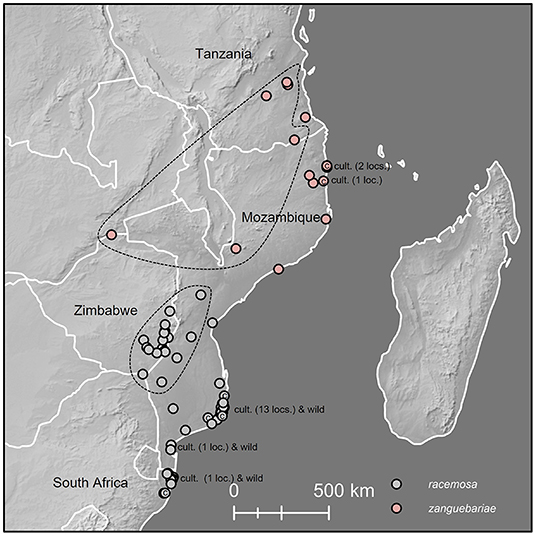
Figure 2. Distribution map for C. racemosa and C. zanguebariae. Dotted lines indicate localities above 150 m for C. racemosa and 200 m for C. zanguebariae. Cult. (cultivated) indicates localities of farmed coffee; loc./locs. = locality/localities.
Bioclim variables (Busby, 1991) from the CHELSA dataset provides modelled climate data at a grid size of 1 km, but these data are modelled and gridded based on local weather stations, which although numerous in some countries are usually spaced many kilometres apart, and are mostly located in urban areas. Depending on local topological features (outcrops, hills, valleys, slope, and aspect), other features of the landscape (rivers, height of the water table, soils, rock outcrops) the local climate and microclimate may differ considerably. Other data sources were therefore used to understand local and microclimate influences (see Fieldwork and Other Observation Data, above).
DNA Sampling, Extraction, Sequencing, and Data Analysis
To assess molecular variation between C. zanguebariae and C. racemosa, we analysed the plastid regions trnL-trnF (trnL intron and trnL-trnF intergenic spacer), rpl16 intron and accD-psaI intergenic spacer, and the nuclear Internal Transcribed Spacer (ITS). These markers have the ability to distinguish between African Coffea species, and identify recently formed hybrids via differential inheritance of plastid and nuclear genomes (Maurin et al., 2007; Davis et al., 2020). In addition to the accessions already available in GenBank for the two species, one for C. zanguebariae and four for C. racemosa (Davis et al., 2020), we analysed five further samples for C. zanguebariae (see Table 1). The C. zanguebariae accession used by Maurin et al. (2007) and Davis et al. (2011) was omitted as its phylogenetic position is ambiguous, most likely due to partial sequences for trnL–F intron, trnL–F intergenic spacer (IGS), and accD–psaI IGS. A final dataset of four accessions for C. racemosa and six accessions for C. zanguebariae was used for illustrative purposes, with C. costatifructa and C. sessiliflora used as outgroups (Table 1). Previous analyses of global and African datasets shows that accessions of C. racemosa (Maurin et al., 2007; Davis et al., 2011; Hamon et al., 2017) and one accession of C. zanguebariae (Davis et al., 2020) belong to a clade of low elevation East African Coffea species, comprising C. costatifructa, C. pocsii, and C. sessiliflora.
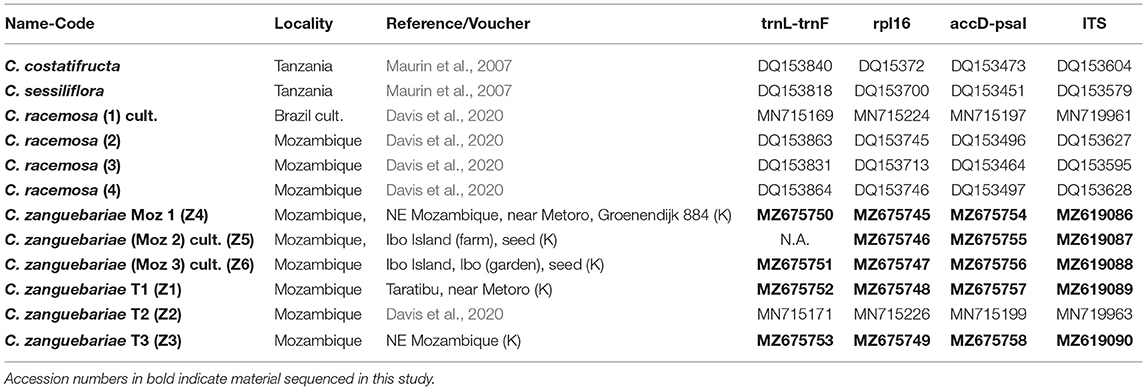
Table 1. List of DNA sampling and accession data.
We extracted DNAs from dried leaves with a modified CTAB protocol (Doyle and Doyle, 1987) and purified them using a QIAquick purification kit (QIAGEN). DNA quality was checked on an agarose gel by electrophoresis. Polymerase chain reactions were conducted as described in Maurin et al. (2007); Sanger sequencing and alignments were carried out following Davis et al. (2020). To reconstruct phylogenetic relationships among the accessions of C. zanguebariae and C. racemosa, the alignments obtained for the plastid and the nuclear regions were concatenated and analysed using Bayesian inference, as implemented in the software MrBayes v.3.2.7 (Huelsenbeck and Ronquist, 2001) with the Beagle library (Ayres et al., 2012). The analysis was carried out using the CIPRES Science Gateway ver. 3.3 (Miller et al., 2010). In MrBayes, we assumed a GTR model of molecular evolution with gamma rate of variation and the sampling parameters were set as following: two runs and four chains for 20,000,000 generations with a relative burn-in of 25%, sampling the chains every 1,000 generations. A maximum clade credibility tree was constructed using FigTree ver. 1.4.4 (Rambaut, 2018).
Flavour Assessment
Several samples of C. racemosa from Hluhluwe (South Africa) and two bulk samples of C. zanguebariae from Ibo Island (Mozambique) were obtained for flavour (aroma and taste) assessment purposes. The C. racemosa samples were supplied by Cultivar Coffee (Hluhluwe Kwa-Zulu Natal, South Africa) a farm specialising in the cultivation of this species; the samples of C. zanguebariae were purchased directly from Ibo Island (northern Mozambique). The C. racemosa samples were wet processed (washed coffee) and carefully processed to optimize quality. The C. zanguebariae samples were sundried locally (on Ibo Island) but were unlikely to have received optimal processing, considering the coffee was not for export but rather for local use.
The samples were sent to two independent coffee professionals and two commercial sensory laboratories, one belonging to a coffee supplier and the other to a roaster and retailer (see Acknowledgements for details). Given that no protocols exist for cupping the two study species, which are inherently very different physically (and possibly chemically) from Arabica and robusta coffee, we asked that the roasting be optimized as best as possible. Simplified or modified versions of the Speciality Coffee Association (SCA) protocol https://sca.coffee/research/protocols-best-practices) were used, with sensory terminology broadly following, where possible, the SCA Coffee Taster's Flavour Wheel (Spencer et al., 2016). No scoring was applied to evaluate the coffees. The aim was not to provide a detailed sensory assessment of each sample and species, but rather to attain a general overview of flavour quality, and cup profile (including flavour notes).
Results
Geographical Distribution and Habitat of C. racemosa and C. zanguebariae
A distribution map for naturally occurring (wild) C. arabica, C. canephora, and wild and cultivated (farmed) C. racemosa and C. zanguebariae is shown in Figure 1; a higher resolution distribution map for C. racemosa and C. zanguebariae is given in Figure 2. The natural distributions of these four species do not overlap.
The indigenous geographical distribution of C. racemosa is central and southern Mozambique, eastern Zimbabwe and northern South Africa (Kwa-Zulu Natal), as reported by Bridson (2003), between latitude 18°20′-28°30′ (Figure 2). Two distinct ecological niches were identified via mapping (including visualization using satellite imagery (GoogleEarth®), herbarium data, and climate profiling (see below): (1) low elevation (<200 m), seasonal (deciduous) coastal forest and bushland, often associated with sandy soils; and (2) low to medium elevation [200–600(−780 m)], seasonal deciduous and semi-evergreen forest, in close proximity to rivers.
Coffea zanguebariae is indigenous in southern Tanzania, northern Zimbabwe and northern Mozambique, between latitude 8°20′-17°30′ (Figure 2). Like C. racemosa, two distinct ecological niches were identified via mapping (including visualization using satellite imagery (GoogleEarth®), herbarium data, and personal observation (see above): (1) low elevation (5–100 m), seasonal (deciduous) coastal forest and bushland, often associated with sandy soils; and (2) low to medium elevation [100–380(−680 m)], seasonal deciduous and semi-evergreen forest, either in close proximity to rivers or amongst rocks or close to rock outcrops. Unlike C. racemosa, the climate profiling analysis (see below) did not identify a distinct bimodality, but this may be due to lower number of ground points, compared to C. racemosa. Herbarium collections made close to the town of Metoro in northern Mozambique (at c. 350 m) did not state habitat information, but direct observation shows that C. zanguebariae grows at the base of large rock outcrops (inselbergs), and near ephemeral water sources in places that are cooler, more humid (and with improved water availability) compared to nearby drier (Miombo) woodland that characterizes the vegetation of the area.
Morphological Review
A summary of the key morphological features of C. racemosa and C. zanguebariae, based on literature (Bridson, 1988, 2003; Burrows et al., 2018), herbarium survey, and fieldwork, is given in Table 2. Some morphological details are illustrated in Figures 3–6. Overall, C. zanguebariae is a larger and more robust plant compared to C. racemosa, being taller (and with a larger stem diameter), with larger leaves, more fruits per inflorescence (and correspondingly per axil and node), and larger seeds. Farmed material of C. racemosa shows distinct variation in seed size and shape (Figures 4B,C). Farmed C. zanguebariae in northern Mozambique shows considerable variation in fruit and seed size (Figures 6A–C); seed size dimorphism in wild C. zanguebariae was reported by Bridson (2003). Both species are deciduous, but may retain their leaves through the dry season in some locations, and in seasons when the dry season is not severe. In farmed conditions, some plants of C. racemosa may lose their leaves, whereas neighbouring plants may not (C. Denison pers. observ.).
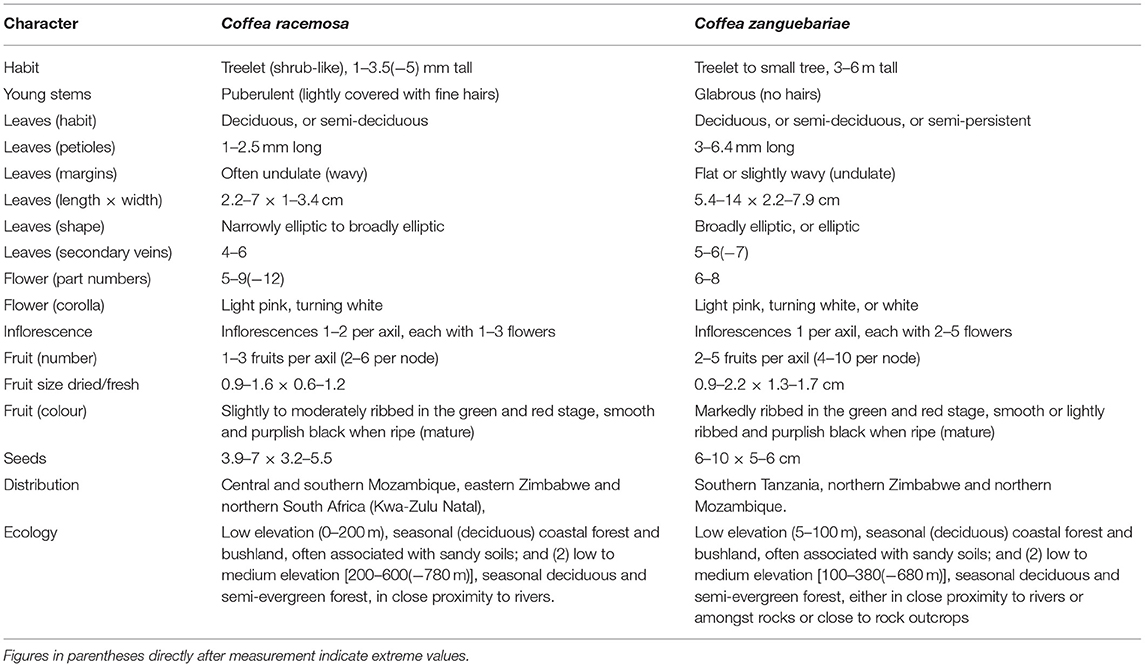
Table 2. Distinguishing morphological characteristics, and distribution and ecology summary for C. racemosa and C. zanguebariae.

Figure 3. Coffea racemosa. (A) Habit of cultivated plant, with immature fruits; (B) Flowers, mature, 5-lobed variant; (C) Fruits, approaching full maturity. Images: Charles Denison.
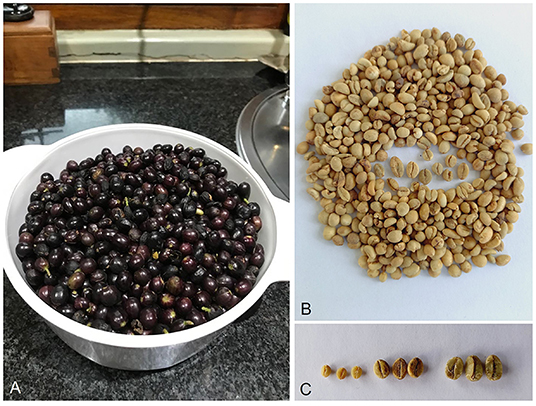
Figure 4. Coffea racemosa. (A) Fresh, fully mature (ripe) fruits; (B) seeds, showing variation in size; (C) seeds (left to right) of Coffea racemosa, compared to C. canephora (robusta) and C. arabica (Arabica). Images: (A) Charles Denison); (B,C) Aaron Davis.

Figure 5. Coffea zanguebariae. (A) Shoots of a cultivated plant, with immature fruits and ripening fruits (red), cultivated near Pema, north-eastern Mozambique; (B) Leaves of a wild plants, for size comparison; (C) Habit of wild plant. (B,C) images taken at Taratibu, near Metoro, near Pemba, north-eastern Mozambique (images: Aaron Davis).

Figure 6. Coffea zanguebariae. (A) Shoots of a cultivated plant, with immature fruits—left, showing upper (adaxial) surface of leaves; right, showing lower (abaxial) surface of leaves; (B) Ripening and almost ripe (mature) fruits, from farmed plants; (C) Fruits, and seeds enclosed in endocarp (parchment)—immediate left, immature fruits (green), uppermost, dried fruits (cherry) and lowermost, parchment coffee, indicating variation in parchment/seed size; (D) unroasted (above) and roasted (below) seeds of C. zanguebariae (four seeds to left), and compared to those of C. racemosa (right). Images (A–C) taken in north-eastern Mozambique, (A,C) on Ibo Island and (B) near Pemba, and (D) in the UK. Images: Aaron Davis.
DNA Analysis
We obtained molecular data for all the plastid and nuclear regions investigated, except the trnL-trnF in one C. zanguebariae sample (M2). Our concatenated plastid matrix included 3,100 characters, and our ITS matrix 792 characters (12 accessions in total). The maximum clade credibility tree resulting from the Bayesian analysis had posterior probabilities (PP) > 0.9 for all clades (Figure 7). A sister relationship for C. racemosa and C. zanguebariae is strongly supported (PP = 1), as are the multiple species-specific samples for each species (PP = 1).
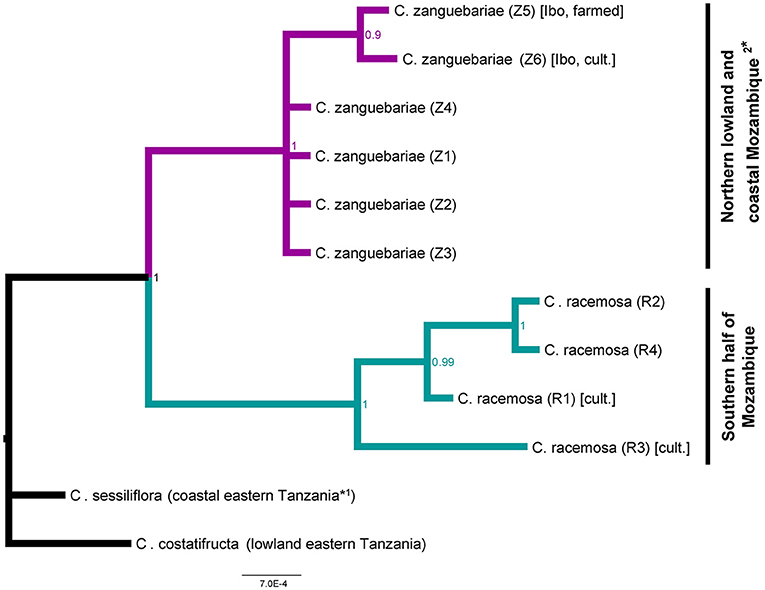
Figure 7. Combined ITS and plastid maximum clade credibility tree. Bayesian posterior probabilities are indicated at nodes. See Table 1 for accession information. Notes: 1* Coffea sessiliflora also occurs in coastal eastern Kenya; 2* C. zanguebariae also occurs in southern Tanzania, and northern Zimbabwe.
Climate Profiling
Our modelled climate data analysis for wild and cultivated C. racemosa gives a mean annual average temperature of 22.9°C and a mean annual precipitation of 807 mm; and for C. zanguebariae 24.8°C and 998 mm. For the wild occurrences of the two main coffee crop species, C. arabica has a mean annual average temperature of 18.7°C and a mean annual precipitation of 1,614 mm, and for C. canephora 23.7°C and 1,596 mm (Table 3). The distribution and density of data points for these two variables, for the four species, are shown in Figures 8, 9. Disaggregated data for cultivated vs. wild for C. racemosa and C. zanguebariae show slightly warmer mean annual temperatures (Bio 1) for cultivated datapoints (0.7 and 1.1°C, respectively), wetter conditions (mean annual precipitation, Bio 12) for cultivated C. racemosa (916 vs. 764 mm) and marginally drier conditions for cultivated C. zanguebariae (969 vs. 990 mm). Rainfall seasonality (Bio 15) is highest (more seasonal) in C. racemosa and C. zanguebariae compared to C. arabica and C. canephora, and there is a second major peak for rainfall seasonality (Bio 15) in C. racemosa, with a mean value midway between C. arabica and C. canephora (Table 3; Figure 9). The lower valued precipitation seasonality (Bio 15) peak for C. racemosa (Figure 4) is due to the lower value returned by coastal datapoints (mean value of 55); the inland points (Figure 2) having a higher precipitation seasonality (mean value 74). Spot checks against publicly available monthly mean precipitation charts for Mozambique and published data (Laíns E Silva, 1954) confirm the distinct precipitation seasonality (i.e., a distinct wet and distinct dry season) for C. racemosa and C. zanguebariae.
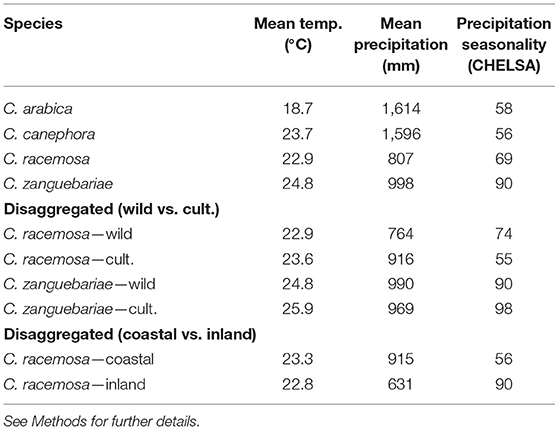
Table 3. Climate profiling summary with mean values and disaggregated mean values for wild and cultivated (C. racemosa and C. zanguebariae), and coastal (low elevation) and inland (higher elevation) for C. racemosa.
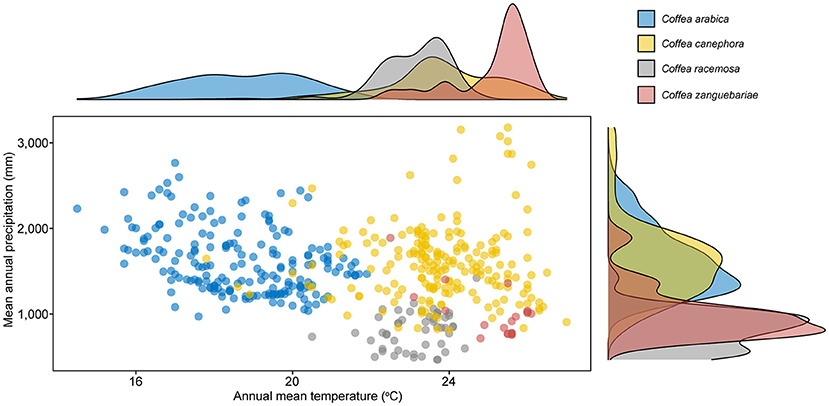
Figure 8. Scatter and density plots of modelled mean annual temperature (CHELSA bio10_01) vs. total mean annual precipitation (CHELSA bio10_012). Mean values in parentheses: Arabica (C. arabica; 18.7°C/1,614 mm); robusta (C. canephora; 23.7°C/1,596 mm); wild and cultivated C. racemosa (22.9°C/807 mm) and wild and cultivated C. zanguebariae (24.8°C/998 mm). Disaggregated values for wild and cultivated (C. racemosa and C. zanguebariae), and coastal (low elevation) and inland (higher elevation) for C. racemosa are given in Table 3. See Methods for further details.
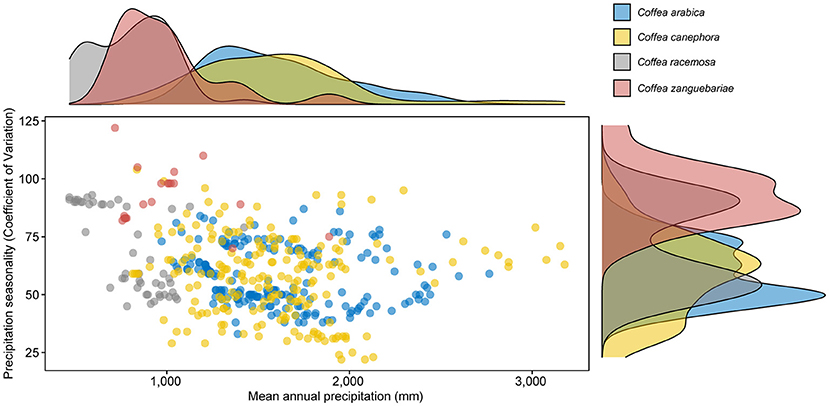
Figure 9. Scatter and density plots of modelled total mean annual precipitation (CHELSA bio10_012) vs. precipitation seasonality (CHELSA bio10_015). Mean values in parentheses: Arabica (C. arabica; 1,614 mm/58); robusta (C. canephora; 1,596 mm); wild and cultivated C. racemosa (807 mm/69) and wild and cultivated C. zanguebariae (998 mm/90). Disaggregated values for wild and cultivated (C. racemosa and C. zanguebariae), and coastal (low elevation) and inland (higher elevation) for C. racemosa are given in Table 3. See Methods for further details.
Farming of C. racemosa and C. zanguebariae
Coffea racemosa is currently being farmed at small scale at Kwa Zulu Natal in South Africa (Hluhluwe) and probably on isolated farms in southern Mozambique; C. zanguebariae is being cultivated on Ibo and Quirimbas Islands, and mainland northern Mozambique (near Pemba). Cultivation of C. zanguebariae on Ibo and Quirimbas Islands (as Ibo coffee) is long-established and probably dates back to at least the 1890s (Cheney, 1925), as reported in the Introduction. Cheney (1925) also mentions, based on an earlier reference (Lanessan, 1886), that in 1880 9,300 kg of “excellent coffee seeds” from C. zanguebariae were harvested on Nossibe (Nosy Bé) an Island off the north-western coast of Madagascar. On reviewing Lanessan (1886), this statement seems to be erroneous, as on page 44 Lanessan states: “In the wild, Coffea zanguebariae Lour. is found on the island, whose grain has a very delicate flavour. Cultivation trials have been made with various varieties of Bourbon coffee (C. arabica) trees, which grow vigorously, even without shelter, and give a high estimated gain in the country. In 1880, 9,300 kg were collected.” And on page 833, Lanessan (1886) states that Coffea zanguebariae grows wild on Nosy Bé'. Clearly then, the assertion made by Cheney (1925) is incorrect, as C. zanguebariae does not grow wild on Nosy Bé, where only a single endemic, wild species is found, viz. C. pervilleana (Davis et al., 2006) and, moreover, Lanessan (1886) is referring to the yield of C. arabica (cv. “Bourbon”) on Nosy Bé, not C. zanguebariae. We were unable to verify whether cultivation of C. zanguebariae (as Zanzibar coffee) still occurs on Zanzibar (Loureiro, 1790; Cheney, 1925), or whether it was brought there from Ibo island, or vice versa, by boat, by Portuguese and other traders, or originated from mainland Tanzania. Given that both Ibo and Zanzibar were key trading ports, a dissemination of cultivated material from island to island is more likely. If we accept the opinion of Loureiro (1790), that C. zanguebariae was taken to Zanzibar by the Portuguese, this species has been cultivated there prior to 1790. The earliest record for the cultivation of C. racemosa is uncertain, although its use as coffee was noted by Hiern (1876) and it is clear from herbarium records (from 1937) that this species was quite widely cultivated in the Inhambane Province of Mozambique (as Inhambane coffee) in the first half of the twentieth century. We have not been able to verify whether cultivation of C. racemosa persists in and around Inhambane, although personal communication with various persons suggest that it is being farmed there on a local scale.
Basic Agronomic Information
The following observation and data records for C. racemosa were made at Hluhluwe, Kwa-Zulu Natal, South Africa, which has a mean annual precipitation of c. 530 mm (with 3 months precipitation of <50 mm, and 6 months <100 mm), and a mean annual temperature of 22.2°C (December, January and February are the hottest months, with mean maximum temperatures of c. 28/29°C). The tree spacing for farmed plots is 1.5 × 2.5 m or 1.5 × 1.5 m. Flowering occurs over a short period from mid-August to mid-September, with flower initiation dependent on the first rains, with fruit development following shortly afterwards. Each flower only lasts about 3 days, and all flowering is over in about 3 weeks. By early November the fruit is at full size, after a rapid ripening and harvest in late November to the end of December of the same year, although small volumes of fruit are harvested in early to mid-January. The period from flowering (anthesis) to harvest (fruit ripening), for farmed plants of this species is thus around 4 months; the same observations are made for wild plants of C. racemosa (Bridson, 2003). At around 7 years old, each (i.e., not fully mature) of the trees under observation produced a mean value of 111 g of ripe fruit (fresh cherry), which converts to c. 12 g of clean, green coffee, giving a conversion rate of 9.4:1 (the farm works on a 10:1 conversion), when using the wet processing method. The mean loss from fresh cherry to pulped coffee (i.e., with the fleshy part removed) is 59.1%; the mean loss from pulped coffee to parchment is 63.9%; and the mean loss from parchment to green is 28.2%.
The following observation and data records for C. zanguebariae were made on Ibo Island, northern Mozambique, which as mean annual precipitation of c. 950 mm (with 7 months precipitation of <50 mm, and a mean annual temperature of 26°C (December, January and February are the hottest months, with mean maximum temperatures of c. 32°C). The tree spacing for farmed plots is 2.5–3 × 2.5–3 m. Flowering occurs over a short period from December to January, with flower initiation dependent on the first major rains. The fruits are almost fully sized by February and March, and harvesting occurs from March to April. On the neighbouring Quirimba island flowering and fruiting times may be 2–4 weeks earlier. The time period from flowering to harvest is around 4 months, as observed for wild plants of C. zanguebariae (Bridson, 1988). Mature trees produce 3–4 kg of fresh cherry, which converts to 600–800 g of clean, green coffee at a conversion rate of 5:1, or 300–400 g (at a 10:1 conversion). Mean losses from fresh to dried cherry, and from dried cherry to clean coffee were not recorded.
Herbarium survey data reveals that C. zanguebariae was probably cultivated in small quantities in central Mozambique at Inhambane, although the specimens for this are incomplete and so their identification is provisional (Bridson, 1988, 2003). We could not find any specimens to support historical reports (Cheney, 1925) of C. zanguebariae being cultivated on Zanzibar (Tanzania) and Ibo Islands (Mozambique), although we have no reason to doubt this. Observation on (and testimony from) Ibo Island, provides evidence of long-established cultivation of C. zanguebriae. Numerous collections from 1937 show that C. racemosa was cultivated on several farms close to and within an 80 km radius of Inhambane. One specimen states that the plants being grown were 35 years old, inferring that C. racemosa had been in cultivation in that region since at least the beginning of the twentieth century. These data generally confirm the historical cultivation information provided by Guerreiro Filho (1992) for these species.
Flavour
All four tasting panels agreed that the C. zanguebariae samples had undergone sub-optimal harvesting, processing, and storage, as evidenced by physical and flavour defects. One sample was notably aged, possibly from the previous seasons harvest. Nevertheless, it was still possible to assess these two samples and make valued judgements on coffee flavour. All four panels remarked that better processed samples of C. zanguebariae would be of interest, and two panels noted that further work on the harvesting and processing of C. racemosa would be worthwhile. All four panels experienced difficulties with roasting due the small size of the beans (seeds). Ideally, a range of roasting levels and profiles would have been preferred, as this has been shown to gain a better understanding of optimal beverage quality in minor coffee crop species and wild coffee species (Davis et al., 2021; B. Bertrand and D. Mieulet pers. comm.). Combined sensory results from the four panels are as follows, but with some of the extremely negative notes (which are assumed to be the result of sub-optimal harvesting, processing, and storage) removed for the two samples of C. zanguebariae.
Coffee racemosa: dry aroma (freshly ground coffee): spice, sweet herbs, liquorice; body: light to medium; acidity: light; flavour notes: blackcurrant, spiced wine, spice, cannabis, star anise, liquorice, buchu (see below), sweet-cake-like, herb-like, cinnamon, cloves, camphor, violet florals, cereal, mint. Buchu, little-known outside South Africa, are plants of the genus Barosma (Rutaceae) and especially Barosma betulina; the leaves and exracted oils are noted for their smell and tasting notes of blackcurrant, spice, and a mixture of rosemary and peppermint.
Coffea zanguebariae: dry aroma: herbal, light eucalyptus, liquorice. Body: light to medium; acidity: light; flavour notes: herbal, savoury, lavender, jasmine, aniseed, liquorice, dark chocolate, spice (cardamon), eucalyptus, medicinal, vanilla, mint. Several cups of coffee were consumed on Ibo Island (A. Davis pers. observ.), including filter, cafetiere and espresso: the quality and flavour was highly variable, but included cups showing considerable promise and remarkable uniqueness.
We found a single example of a flavour appraisal of C. racemosa by a coffee roaster/retailer (https://www.coffeereview.com/review/south-africa-elephant-coast-estate-coffea-racemosa/). On the basis of a blind tasting, following a medium-light roast of coffee purchased from South Africa (Hluhluwe), their assessment notes are as follows (with some minor editing): richly bittersweet, deep-toned; flavour notes: hop flowers, pink peppercorn, tangerine zest, quince, fresh-cut fir in aroma and cup; sweet-savoury structure with friendly, accessible acidity; satiny-smooth mouthfeel; the bittersweet finish centres around tangerine zest and quince; basically sweet, this coffee's main impulses are a unique and intriguing citrus-zest-like bittersweetness with savoury floral underpinnings.
Discussion
Species Status and Systematic Affinities
Our DNA analyses infer that C. racemosa and C. zanguebariae are two closely related but separate entities, which given their morphological (Table 1) and molecular (Figure 7) dissimilarity should be recognized at the level of species, supporting the taxonomic conclusions of Bridson (1988, 2003). Based on our work and previous molecular analyses (Maurin et al., 2007; Davis et al., 2011, 2020; Hamon et al., 2017) we know that these two species belong to a well-supported clade of low to mid elevation East African Coffea species (Davis et al., 2006), comprising C. costatifructa (eastern Tanzania; 10–700 m), C. pocsii (eastern Tanzania; 270–600 m), C. racemosa (eastern Zimbabwe, Mozambique, South Africa (Kwa Zulu Natal; 0–600 m), C. sessiliflora (south-eastern Kenya and eastern Tanzania; 10–500 m) and C. zanguebariae [southeaster Tanzania, north-eastern Mozambique; 10–350(−600) m].
Based on taxonomic literature review (Bridson, 1988, 2003), herbarium specimen observation and field work for C. racemosa and C. zanguebariae, it is clear that species can be easily distinguished morphologically (Table 1). Overall, C. zanguebariae is a larger and more robust plant compared to C. racemosa, being taller (and with a larger stem diameter), with larger leaves, more fruits per inflorescence (and correspondingly for each axil and node), and larger fruits and seeds. The shoots of C. zanguebariae lack the fine covering of short hairs evident on the shoots of C. racemosa, which is a useful criterion for differentiating between sterile plants (i.e., flowers or fruits absent), and even sterile plants lacking leaves, (i.e., during the dry season).
With the species relationships for these species resolved, and with additional knowledge from literature review and field observation, it is possible to confidently assign common (vernacular) names to these species. The vernacular names for C. racemosa include: Inhambane coffee; and for C. zanguebariae: Ibo coffee and Zanzibar coffee.
Climate Profiling
The climate profiling analysis returned a mean annual average temperature and a mean annual precipitation of 22.9°C/807 mm for C. racemosa (wild and cultivated), and 24.8°C/ 998 mm C. zanguebariae (wild and cultivated). Disaggregated values for wild and cultivated occurrences are given in Table 3. For the indigenous occurrences of the two main coffee crop species, C. arabica returned values of 18.7°C and 1,614 mm, and for C. canephora 23.7°C and 1,596 mm (Table 3; Davis et al., 2021). These data support assumptions and reports of tolerance to high temperatures and low precipitation (Laíns E Silva, 1954; Krug, 1965; Halle and Faria, 1973; Guerreiro Filho, 1992), which would, of course, be expected given the natural distribution of these two species. The higher precipitation values accorded to these species by Krug (1965) and as also reported in Guerreiro Filho (1992), i.e., of 1,300 mm and 1,600 mm, are considered erroneous; these amounts being similar to those experienced by indigenous and cultivated C. arabica and C. canephora, which occur natutrally in much wetter areas of Africa.
Other climate parameters are of note, including seasonality, and mean absolute maximum temperatures. Review of publicly available climate charts and published data (Laíns E Silva, 1954; Guerreiro Filho, 1992) shows that both species come from distinctly seasonal environments, with lengthy periods of low monthly rainfall (see also Basic Agronomic Information). In the Inhambane Province in Mozambique, i.e., location of wild and cultivated C. racemosa, data from five lowland (<100 m) climate stations shows this species can experience 3–7 months precipitation of <50 mm, and 8–10 months with <100 mm) per annum; the values for the mean annual precipitation were reported between as 682–1,113 mm (Laíns E Silva, 1954). For the same five localities, the following temperatures ranges were reported (Laíns E Silva, 1954): mean maximum temperatures 28.8–30.3°C; mean minimum 9–18.4°C; absolute maximum 33–46°C; and absolute minimum 3–14°C. Temperature ranges for C. zanguebariae are likely to be similar, or somewhat warmer based on the modelled mean annual temperature provided by our climate profile analysis. Ad hoc absolute maximum temperatures (in shade) of 39–45°C were recorded in situ in January and February 2019 (A. Davis pers. observ.) on Ibo and Quirimbas islands in northern Mozambique, using a portable thermo-hygrometer (Tinytag® TH-2500), plants growing nearby were either in shade, partial shade or full sun. Shaded plants appeared to be in better health, had larger leaves, more exuberant growth and more fruits.
Laíns E Silva (1954) remarked that it was surprising that the farming of C. racemosa had not been attempted in the Mediterranean region of Europe, given the climate data reported for this species, although mean and absolute low temperatures would probably pose the most obvious constraint to successful cultivation.
Key features for adaptation to this warm, low precipitation, and seasonal environment include deciduous habit, and the short period of fruit development of around 4 months, from flower (i.e., anthesis) to mature (harvestable) fruit. Other key traits are not clearly apparent but may include the short flowering period (individual flowers and total flowering period per plant) and perhaps mature fruit colour (purplish to purple black). Root characteristics (e.g., type, dimensions, architecture, microbiome) may also play a role. Further research in this area would be worthwhile, as would more direct measurement of above-ground (e.g., temperatures, relative humidity, vapour pressure deficit) and below-ground (soil moisture, soil water potential, temperature) climate metrics.
The rainfall bimodality revealed in the climate profiling analysis for precipitation seasonality for C. racemosa (Table 3; Figure 9) corresponds to the distribution (Figure 2) and ecological differences between coastal (< 200 m) and inland populations [200–600(−780 m)]. Whilst this may be interpreted as climate partitioning, with the higher elevation populations experiencing slightly lower mean annual temperature (Bio 1), lower precipitation (Bio 12) and higher precipitation seasonality (Bio 15), as shown in Table 3, it is likely the precipitation parameters are offset by the availability of ground water from nearby or adjacent rivers and local environmental features (e.g., adjacency to large rock outcrops) and other microhabitats with higher levels of soil moisture).
Agronomic and Value Chain Considerations
Yields are low for C. racemosa and C. zanguebariae, compared to the mainstream crop species (C. arabica and C. canephora). Yields of C. zanguebariae appears to be higher than C. racemosa, with yields estimated between 300 and 400 g of clean coffee per plant, based on a conversion ratio of 10:1; a planting density of 1,000 plants per hectare would provide 300–400 kg/ha of clean coffee. This is equivalent to lower yield rates of Arabica coffee. At the same conversion ratio and planting density, the yield of C. racemosa reported here would give 111 kg/ha, representing a very low yield for commercial coffee, although this is for young and perhaps not fully mature plants. Further yield experimentation is required for both species.
The small seed size of C. racemosa and C. zanguebariae (Table 2; Figures 4, 6) is problematic, due to its contribution to reduced yields, and issues concerned with processing. Small seeds could also be problematic for roasting, as they may require specific equipment and a roasting profile that differs considerably from the main crop species (Arabica and robusta).
Agronomic traits of obvious value for C. racemosa and C. zanguebariae are extreme heat tolerance, low precipitation tolerance, and extreme precipitation seasonality, i.e., long dry season (number of months with low precipitation values) (Table 3; Figures 8, 9). Such attributes would be key to the success of a coffee crop in areas where use of the main production species would be climatically unviable, and could be useful for the production of drought tolerant coffee crop cultivars via interspecies breeding. Indeed, C. racemosa has been successfully crossed with C. arabica (and backcrossed with C. arabica) to produce a range of hybrid derived cultivars, notably C. arabica cv. ‘Siriema' (sometimes referred to more broadly as C. ‘Aramosa') although crosses have also been made with C. canephora (Sureshkumar et al., 2010), and the interspecies hybrid C. × ‘Congusta' (C. congensis × C. canephora) (Sureshkumar et al., 2004). These hybrids have been shown to demonstrate a degree of drought tolerance, compared to C. arabica (Grisi et al., 2008; Melo et al., 2014; Carvalho et al., 2017) and C. canephora (Sureshkumar et al., 2004).
The extremely short fruit ripening period [i.e., the duration from flowering (anthesis) and full fruit ripeness] of around 4 months represent another key trait for C. racemosa and C. zanguebariae, which has already been investigated by workers. Early ripening plants have been reported for C. arabica × C. racemosa hybrids, with a 180 day (around 6 months) fruit ripening period, as opposed to a 6–8 month period in C. arabica (Sondahl et al., 1997). In hybrids of C. canephora × C. racemosa a 160–170 (around five and a half months) fruit ripening period has been reported (Sureshkumar et al., 2004, 2010), as opposed to 9–11 months in C. canephora.
Originally, the objective of producing hybrids of C. arabica × C. racemosa was to introduce resistance to coffee leaf miner (Perileucoptera coffeella) (Medina Filho et al., 1977b), although C. racemosa has also been trialled in breeding work to reduce the caffeine content in C. arabica (Medina Filho et al., 1977a).
Flavour
The flavour (aroma and taste) of C. racemosa and C. zanguebariae is conspicuously distinct from C. arabica (Arabica) and C. canephora (robusta), mainly due to the presence of unique flavour notes. In particular, many volatile-herbal (e.g., eucalyptus, lavender mint, fir tree), and spice (camphor, cinnamon) notes, and savoury elements immediately set this apart from the regular coffee flavour experience. Many of the tasting notes identified (e.g., jasmine, cinnamon, chocolate, vanilla, blackcurrant) would be accepted as positive flavour attributes for coffee (Spencer et al., 2016); others (e.g., camphor, eucalyptus, savoury, medicinal) probably would be considered as unusual, challenging, and perhaps undesireable. Without doubt, C. racemosa and C. zanguebariae considerably broaden the sensory envelope of coffee drinking.
Conclusion
In this contribution we demonstrated that C. racemosa (Inhambane coffee) and C. zanguebariae (Ibo coffee, or Zanzibar coffee) are two closely related (sister) species, which are easily differentiated from each other on the basis of their morphology, DNA, and distribution; and to some extent climate profile (annual mean temperature) and flavour. Both species experience high temperatures, low precipitation (rainfall), and distinct precipitation seasonality (a long dry season), in their natural and cultivated environments in East Africa. With a cultivation history of over 200 years, these species are farmed at the distinctly local scale in Mozambique and South Africa, although they may have been grown at greater scale and frequency up until the middle of the last century. Today, they probably represent the world's rarest production coffee species, although many wild Coffea species are gathered locally for use as coffee (Davis et al., 2019).
Coffea racemosa and C. zanguebariae possess useful traits for coffee crop development, especially under accelerated climate change, due to their tolerance of hight temperatures, low total annual precipitation, and marked precipitation seasonality (long dry season), and short fruit ripening period (around 4 months). In terms of flavour, C. racemosa and C. zanguebariae are unique, and substantially broaden the sensory experience of coffee drinking. Negative traits for these species include low yield, and small seed (coffee bean) size, and, for some perhaps, their unique flavour notes. Further experimentation is required to more fully understand the climate tolerances of these species, particularly in situ, and to understand the underlying mechanisms of adaptation. Further work on sensory assessment, seed (coffee bean) chemistry, and agronomy, is also required.
Coffea racemosa has already been utilized in breeding programmes via interspecies hybridization, for pest resistance and drought tolerance, and shortening the duration of fruit development and ripening. Coffea zanguebariae has not been utilized in this way, probably due to confusion with other species (e.g., C. racemosa and C. pseudozanguebariae). The greater robustness (plant and leaf size), higher yield, and larger seed size may provide additional useful traits. The considerable morphological variation of C. zanguebariae and C. racemosa, particularly for fruit and seed size, and possible flavour diversity, suggests that they would benefit from pre-farm and farm selection for within-species crop improvement.
Data Availability Statement
The data presented in the study are deposited in the Genbank DNA sequence repository, accession numbers are provided in Table 1.
Author Contributions
AD, JM, and IA devised the overall framework of the study. AD undertook the literature survey and herbarium data collection, participated in fieldwork, and was the lead on writing the manuscript. RG undertook the DNA sequencing and analyses and contributed toward the writing of the manuscript. JM led on the climate profiling analyses and contributed toward the writing of the manuscript. AD, IA, and MC undertook the fieldwork and farm survey in Mozambique and CD in South Africa. All the authors devised the elements of the study and contributed data and manuscript refinement.
Funding
We are grateful to the Bentham-Moxon Trust (Charity Number: 305966) for funding fieldwork in Mozambique, and the Amar-Franses and Foster-Jenkins Trust for supporting the purchase of equipment and other research costs.
Conflict of Interest
CD operates a small farm producing C. racemosa in Hluhluwe (South Africa), from where agronomic data was recorded.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We gratefully acknowledge the support and collaboration of the following organizations in Mozambique: University of Lúrio (Pemba), Cabo Delgado Provincial Directorate of Culture and Tourism, and Associação Fruticultores e Horticultores de Cabo Delgado (FRUTICAD). We would like to thank individuals in Mozambique for assisting us in our research: Issa Tarmamade (Ibo District's Administrator), Ruben Morgado (Fundação Carlos Morgado), José Soares (MozFIP, Maputo), Luis Augusto (FRUTICAD) and Madyo Couto (MozBio, Maputo). Elsewhere we would like to thank: the directors and staff at the Royal Botanic Gardens, Kew (K); Natural History Museum, London (BM); Muséum National d'Histoire Naturelle, Paris (P); National Botanic Garden of Belgium, Meise (BR); and National Herbarium Nederland, Wageningen (WAG), for access to herbarium collections; and staff at Union Hand-Roasted Coffee (London) and staff at WR Wakefield (London), James Hoffmann (Square Mile Coffee Roasters, London), and Tim Wendelboe (Tim Wendelboe AS, Oslo) for providing coffee tasting (flavour) assessments.
References
Ayres, D. L., Darling, A., Zwickl, D. J., Beerli, P., Holder, M. T., Lewis, P. O., et al. (2012). BEAGLE: an application programming interface and high-performance computing library for statistical phylogenetics. Syst. Biol. 61, 170–173. doi: 10.1093/sysbio/syr100
Bridson, D. M. (1982). Studies in Coffea and Psilanthus for part 2 of 'Flora of Tropical East Africa': Rubiaceae. Kew Bull. 36, 817–860. doi: 10.2307/4117921
Bridson, D. M. (1988). “Coffea,” in Flora of Tropical East Africa, Rubiaceae, eds R. M. Polhill, D. M. Bridson and B. Verdcourt. (Rotterdam/Brookfield: Balkema), 703–723.
Bridson, D. M. (2003). “Coffea,” in Flora Zambesiaca, ed G. V. Pope (Kew: Royal Botanic Gardens), 452–463.
Burrows, J., Burrows, S., Lötter, M., and Schmidt, E. (2018). Trees and Shrubs of Mozambique. South Africa: Jacana.
Busby, J. R. (1991). “BIOCLIM—a bioclimatic analysis and prediction system,” in Nature Conservation: Cost Effective Biological Surveys and Data Analysis, eds C.R. Margules and M.P. Austin (Canberra: Commonwealth Scientific and Industrial Research Organisation), 64–68.
Carvalho, F. G., Sera, G. H., Andreazi, E., Sera, T., De Barista Fonseca, I. C. B., Carducci, C., et al. (2017). Drought tolerance in seedlings of coffee genotypes carrying genes of different species. Coffee Sci. 12, 9–16. doi: 10.25186/cs.v12i2.1175
Cheney, R. H. (1925). A Monograph of the Economic Species of the Genus Coffea L. New York, NY: The New York University Press.
Chevalier, A. (1942). Les caféiers du globe, fasc. 1: Iconographie des caféiers sauvages et cultivés. Ency. Biol. 22, 1–36.
Chevalier, A. (1947). Les caféiers du globe, fasc. 3: systématique des caféiers et faux-caféiers. Maladies et insectes nuisibles. Ency. Biol. 28:212.
Cramer, P. J. S. (1957). A Review of Literature of Coffee Research in Indonesia. Turrialba: SIC Editorial, Inter-American Institute of Agricultural Sciences.
DaMatta, F. M. (2004). Exploring drought tolerance in coffee: a physiological approach with some insights for plant breeding. Braz. J. Plant. Physiol. 16, 1–6. doi: 10.1590/S1677-04202004000100001
DaMatta, F. M., and Ramalho, J. D. C. (2006). Impacts of drought and temperature stress on coffee physiology and production: a review. Braz. J. Plant. Physiol. 18, 55–81. doi: 10.1590/S1677-04202006000100006
Davis, A. P., Chadburn, H., Moat, J., O'sullivan, R., Hargreaves, S., and Nic Lughadha, E. (2019). High extinction risk for wild coffee species and implications for coffee sector sustainability. Sci. Adv. 5:eaav3473. doi: 10.1126/sciadv.aav3473
Davis, A. P., Gargiulo, R., Fay, M. F., Sarmu, D., and Haggar, J. (2020). Lost and found: Coffea stenophylla and C. affinis, the forgotten coffee crop species of West Africa. Front. Plant Sci. 11:616. doi: 10.3389/fpls.2020.00616
Davis, A. P., Govaerts, R., Bridson, D. M., and Stoffelen, P. (2006). An annotated taxonomic conspectus of the genus Coffea (Rubiaceae). Bot. J. Linn. Soc. 152, 465–512. doi: 10.1111/j.1095-8339.2006.00584.x
Davis, A. P., Mieulet, D., Moat, J., Sarmu, D., and Haggar, J. (2021). Arabica-like flavour in a heat-tolerant wild coffee species. Nat. Plants 7, 413–418. doi: 10.1038/s41477-021-00891-4
Davis, A. P., and Rakotonasolo, F. (2021). Six new species of Coffea from northern Madagascar. Kew Bull. 76:5. doi: 10.1007/s12225-021-09952-5
Davis, A. P., Tosh, J., Ruch, N., and Fay, M. F. (2011). Growing coffee: Psilanthus (Rubiaceae) subsumed on the basis of molecular and morphological data; implications for the size, morphology, distribution and evolutionary history of Coffea. Bot. J. Linn. Soc. 167, 357–377. doi: 10.1111/j.1095-8339.2011.01177.x
Davis, A. P., Wilkinson, T., Challa, Z. K., Williams, J., Baena, S., Gole, T. W., and Moat, J. (2018). Coffee Atlas of Ethiopia. London: Kew Publishing.
Doyle, J. J., and Doyle, J. L. (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phyt. Bull. Bot. Soc. Amer. 19, 11–15.
Grisi, F. A., Donizeti, J., De Castro, E. M., De Oliveira, C., Biagiotti, G., and De Melo, L. A. (2008). Leaf anatomical evaluations in 'catuaí' and “siriema” coffee seedlings submitted to water stress. Ciênc. Agrotec. 32, 1730–1736. doi: 10.1590/S1413-70542008000600008
Hamon, P., Grover, C. E., Davis, A. P., Rakotomalala, J.-J., Raharimalala, N. E., Albert, V. E., et al. (2017). Genotyping-by-sequencing provides the first well-resolved phylogeny for coffee (Coffea) and insights into the evolution of caffeine content in its species. Mol. Phylogenet. Evol. 109, 351–361. doi: 10.1016/j.ympev.2017.02.009
Hiern, W. P. (1876). On the African species of the genus Coffea. Trans. Linn. Soc. London Bot. 1, 169–176. doi: 10.1111/j.1095-8339.1876.tb00033.x
Holmgren, P. K., Holmgren, N. H., and Barnett, L. C. (1990). Index Herbariorum. Part 1: The Herbaria of the World, 8th edn. Regnum Vegetabile.: New York, NY: Botanical Garden.
Huelsenbeck, J. P., and Ronquist, F. (2001). MrBAYES: Bayesian inferenceof phylogenetic trees. Bioinformatics 17, 754–755. doi: 10.1093/bioinformatics/17.8.754
IPCC (2014). “Climate change 2014: synthesis report,” in Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds Core Writing Team, R.K. Pachauri and L.A. Meyer (Geneva: IPCC), 151.
Karger, D. N., Conrad, O., Böhner, J., Kawohl, T., Kreft, H., Soria-Auza, R. W., et al. (2017). Climatologies at high resolution for the earth's land surface areas. Sci. Data 4:170122. doi: 10.1038/sdata.2017.122
Kassambara, A. (2020). ggpubr: 'ggplot2' Based Publication Ready Plots. R package version 0.4.0. Availale online at: https://CRAN.R-project.org/package=ggpubr
Kath, J., Byrareddy, V. M., Craparo, A., Nguyen-Huy, T., Mushtaq, S., Cao, L., et al. (2020). Not so robust: Robusta coffee production is highly sensitive to temperature. Glob. Chang. Biol. 26, 3677–3688. doi: 10.1111/gcb.15097
Laíns E Silva, H. J. (1954). Subsídios Para o Fomento da Cultura do Café em Moçambique. Agência Geral do Ultramar. Lisboa: Divisão de Publicações e Biblioteca.
Lanessan, J. L. (1886). Les Plantes Utiles des Colonies Françaises. Imprimerie Nationale. doi: 10.5962/bhl.title.61300
Lopes, M. H. C. (1974). Caractéristiques chimiques et technologiques des cafés Racemosa du Mozambique. Café Cacao Thé 18, 263–276.
Maurin, O., Davis, A. P., Chester, M., Mvungi, E. F., Jaufeerally-Fakim, Y., and Fay, M. F. (2007). Towards a phylogeny for Coffea (Rubiaceae): identifying well-supported lineages based on nuclear and plastid DNA sequences. Ann. Bot. 100, 1565–1583. doi: 10.1093/aob/mcm257
Medina Filho, H. P., Carvalho, A., and Medina, D. M. (1977a). Germoplasma de C. racemosa e seu potencial no melhoramento do cafeeiro. Bragantia 36, 43–46. doi: 10.1590/S0006-87051977000100040
Medina Filho, H. P., Carvalho, A., and Monaco, L. (1977b). Melhoramento do cafeeiro. XXXVII—Observações sobre a resistência do cafeeiro ao bicho mineiro. Bragantia 36, 131–137. doi: 10.1590/S0006-87051977000100011
Melo, E. F., Fernandes-Brum, C. N., Pereira, F. J., De Castro, E. M., and Chalfun-Júnior, A. (2014). Anatomic and physiological modifications in seedlings of Coffea arabica cultivar Siriema under drought conditions. Ciênc. Agrotec. 38, 25–33. doi: 10.1590/S1413-70542014000100003
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2010). “Creating the CIPRES Science Gateway for inference of large phylogenetic trees,” in Proceedings of the Gateway Computing Environments Workshop (GCE), (New Orleans, LA), 1–8.
Moat, J., Gole, T. W., and Davis, A. P. (2019). Least Concern to Endangered: applying climate change projections profoundly influences the extinction risk assessment for wild Arabica coffee. Glob. Chang. Biol. 25, 390–403. doi: 10.1111/gcb.14341
Moat, J., Williams, J., Baena, S. T. W., Gole, T. W., et al. (2017). Resilience potential of the Ethiopian coffee sector under climate change. Nat. Plants 3:17081. doi: 10.1038/nplants,0.2017.81
R Core Team (2016). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Rambaut, A. (2018). FigTree, version 1.4.4. Available onine at: http://github.com/rambaut/figtree/ (accessed July 11, 2020).
Silva, H. L. (1956). Número de cromossamas en Coffea racemosa Lour. Bragantia 15, 17–18. doi: 10.1590/S0006-87051956000100034
Sondahl, M. R., Petraco, M., and Zambolim, L. (1997). “Breeding for qualittative traits in Arabica coffee,” in Prodeedings of the 10th Colleque Scientifique seu le Café (Nairobi, Kenta), 447–456.
Spencer, M., Sage, E., Velez, M., and Guinard, J.-X. (2016). Using single free sorting and multivariate exploratory methods to design a new coffee taster's flavor wheel. J. Food Sci. 81, S2997–S3005. doi: 10.1111/1750-3841.13555
Sureshkumar, V. B., Prakash, N. S., and Mohanan, K. V. (2010). A study of Coffea racemosa × Coffea canephora var. robusta hybrids in relation to certain critically important characters. Int. J. Plant Breed. Genet. 4, 30–35. doi: 10.3923/ijpbg.2010.30.35
Sureshkumar, V. B., Selvakumar, M., Santaram, A., and Devasia, J. (2004). “Performance evaluation of interspecific hybrids of Coffea racemosa and Sln. 35 (Coffea congensis × Coffea canephora) under Indian conditions,” in Proceedings of the 20th International Conference on Coffee Science.
Thiers, B. (2019). Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York, NY: Botanical Garden's Virtual Herbarium. Available online at: http://sweetgum.nybg.org/science/ih/ (accessed September 11, 2019).
Keywords: climate change, coffee, DNA sequencing, climate profiling, coffee agronomy, plastid (chloroplast) DNA, Internal Transcribed Spacer (ITS)
Citation: Davis AP, Gargiulo R, Almeida INdM, Caravela MI, Denison C and Moat J (2021) Hot Coffee: The Identity, Climate Profiles, Agronomy, and Beverage Characteristics of Coffea racemosa and C. zanguebariae. Front. Sustain. Food Syst. 5:740137. doi: 10.3389/fsufs.2021.740137
Received: 12 July 2021; Accepted: 23 September 2021;
Published: 21 October 2021.
Edited by:
Paulo Mazzafera, State University of Campinas, BrazilReviewed by:
Benoit Bertrand, Institut National de la Recherche Agronomique (INRA), FranceAejaz Ahmad Dar, Sher-e-Kashmir University of Agricultural Sciences and Technology of Jammu, India
Copyright © 2021 Davis, Gargiulo, Almeida, Caravela, Denison and Moat. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aaron P. Davis, a.davis@kew.org