
Isolation, Characterization and In Silico Studies of Secondary Metabolites from the Whole Plant of Polygala inexpectata Peşmen & Erik


 , , ,
, , ,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Isolation
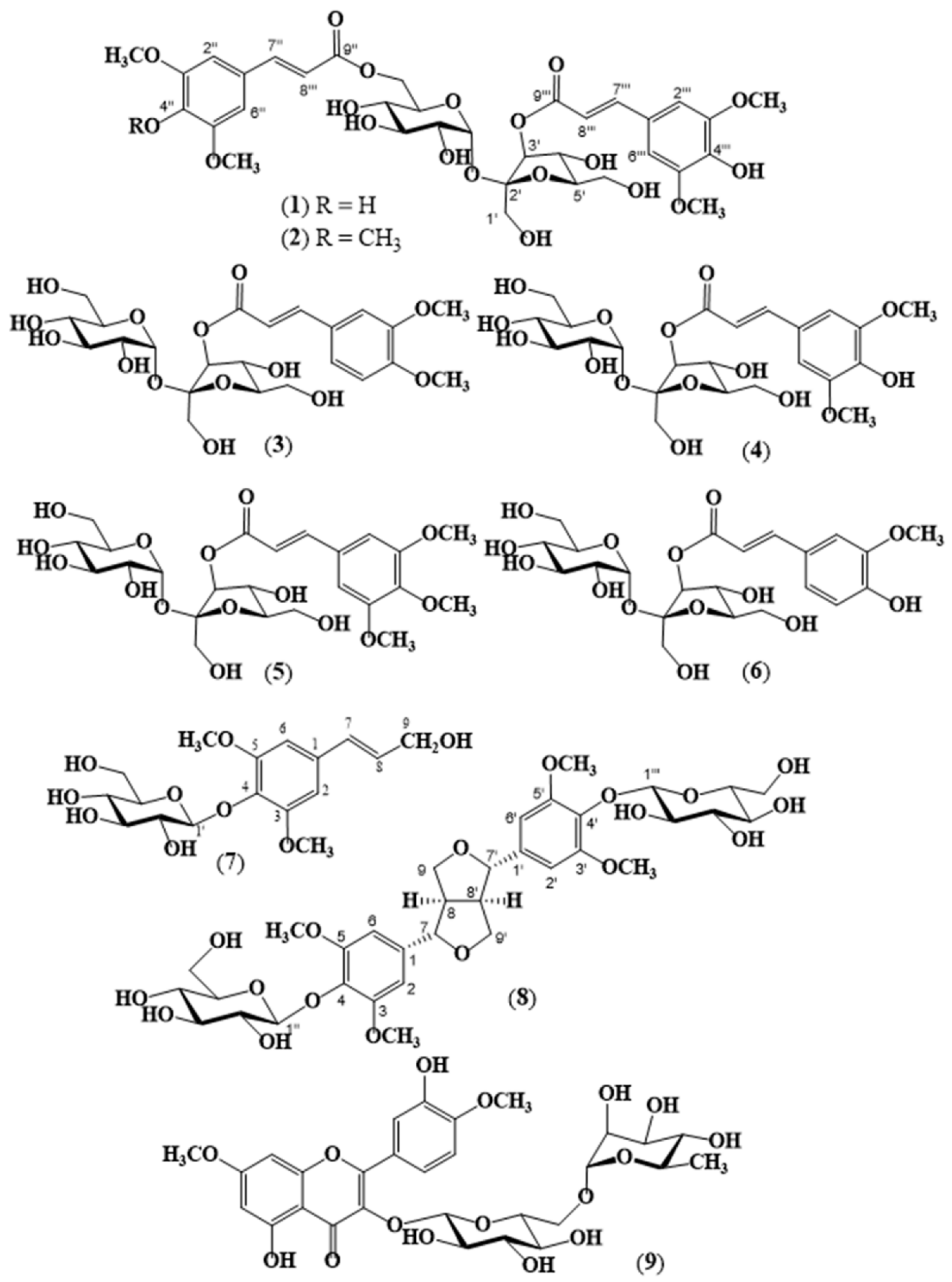
4.4. Physical and Spectral Data of Isolated Compounds 1 to 9
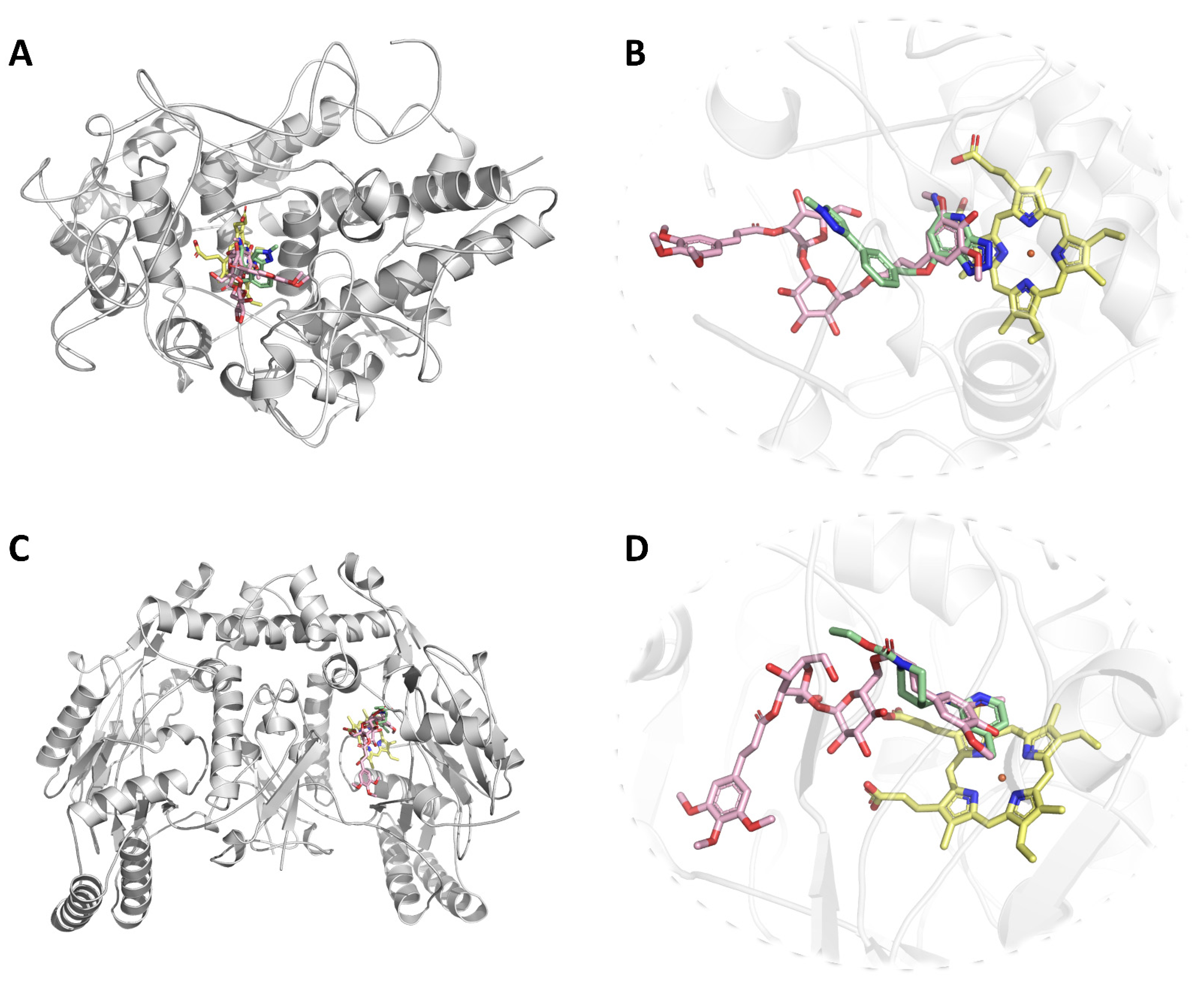
4.5. Protein-Ligand Docking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cronquist, A. Polygalaceae. In An Integrated System of Classification of Flowering Plants; Columbia University Press: New York, NY, USA, 1981; pp. 775–778. [Google Scholar]
- Paiva, J.A.R.; De Cuba, A.D.C. Polygalarum africanarum et madagascariensium prodromus atque gerontogaei generis Heterosamara Kuntze, a genere Polygala L. segregati et a nobis denuo recepti, synopsis monographica. Cyanus 1998, 1, 346. [Google Scholar]
- Bernardi, L.F. Consideraciones taxonomicaly fitogeograficas acerca de 101 Polygalae americanas. Cavanillesia Altera 2000, 1, 1–456. [Google Scholar]
- Cullen, J. Polygala L. In Flora of Turkey and the East Aegean Islands, 1st ed.; Davis, P.H., Ed.; Edinburgh University Press: Edinburgh, Scotland, 1965; pp. 533–539. [Google Scholar]
- Peşmen, H. Six new species from Anatolia. Roy. Bot. Gard. 1980, 38, 435–441. [Google Scholar]
- Davis, P.H.; Mill, R.R.; Tan, K. Polygala L. Flora of Turkey and the East Aegean Islands, 10th ed.; Edinburgh University Press: Edinburgh, Scotland, 1988; pp. 64–65. [Google Scholar]
- Eren, Ö.; Parolly, G.; Raus, T.; Kürschner, H. A new species of Polygala L. (Polygalaceae) from south-west Anatolia. Bot. J. Linn. Soc. 2008, 158, 82–86. [Google Scholar] [CrossRef]
- Dönmez, A.A.; Uğurlu Aydın, Z.; Işık, S. Polygala turcica (Polygalaceae), a new species from E Turkey, and a new identification key to Turkish Polygala. Willdenowia 2015, 45, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Dönmez, A.A.; Uğurlu Aydın, Z. Polygala azizsancarii (Polygalaceae), a new species from Mardin Province, SE Turkey. Phytotaxa 2018, 340, 255–262. [Google Scholar] [CrossRef]
- Baytop, T. Türkiye’de Bitkiler ile Tedavi, Geçmişte ve Bugün; Nobel Tıp Kitabevleri: İstanbul, Turkey, 1999. [Google Scholar]
- Ban, J.Y.; Lee, H.J.; Lee, S.B.; Lee, Y.J.; Seong, N.S.; Song, K.S.; Seong, Y.H. Methanol extract of Polygalae radix protects excitotoxicity in cultured neuronal cells. Korean J. Med. Crop. Sci. 2003, 11, 298–305. [Google Scholar]
- Suksri, S.; Premcharoen, S.; Thawatphan, C.; Sangthongprow, S. Ethnobotany in Bung Khong Long non-hunting area, northeast Thailand. Kasetsart J. (Nat. Sci.) 2005, 39, 519–533. [Google Scholar]
- Lin, L.L.; Huang, F.; Chen, S.B.; Yang, D.J.; Chen, S.L.; Yang, J.S.; Xiao, P.G. Xanthones from the roots of Polygala caudata and their antioxidation and vasodilatation activities in vitro. Planta Med. 2005, 71, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Li, T.Z.; Zhang, W.D.; Yang, G.J.; Liu, W.Y.; Liu, R.H.; Zhang, C.; Chen, H.S. New flavonol glycosides and new xanthone from Polygala japonica. J. Asian. Nat. Prod. Res. 2006, 8, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Alagammal, M.; Lincy, M.P.; Mohan, V.R. Hepatoprotective and Antioxidant effect of Polygala rosmarinifolia Wight & Arn against CCl4 induced hepatotoxicity in rats. J. Pharmacogn. Phytochem. 2013, 2, 118–124. [Google Scholar]
- Wu, G.; Gao, Y.; Kang, D.; Huang, B.; Huo, Z.; Liu, H.; Liu, X. Design, synthesis and biological evaluation of tacrine-1, 2, 3-triazole derivatives as potent cholinesterase inhibitors. Med. Chem. Comm. 2018, 9, 149–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Cui, Y.; Wu, P.; Zhao, P.; Zhou, Q.; Zhang, Z.; Zhang, X. Polygalae radix: A review of its traditional uses, phytochemistry, pharmacology, toxicology, and pharmacokinetics. Fitoterapia 2020, 104759. [Google Scholar] [CrossRef]
- Ikeya, Y.; Sugama, K.; Okada, M.; Mitsuhashi, H. Two xanthones from Polygala tenuifolia. Phytochemistry 1991, 30, 2061–2065. [Google Scholar] [CrossRef]
- Fujita, T.; Da-You, L.; Ueda, S.; Takeda, Y. Xanthones from Polygala tenuifolia. Phytochemistry 1992, 31, 3997–4000. [Google Scholar] [CrossRef]
- Ikeya, Y.; Sugama, K.; Maruno, M. Xanthone C-glycoside and acylated sugar from Polygala tenuifolia. Chem. Pharm. Bull. 1994, 42, 2305–2308. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, T.R.; Cechinel Filho, V.; Santos, A.R.; Calixto, J.B.; Delle Monache, F.; Pizzolatti, M.G.; Yunes, R.A. Three xanthones from Polygala cyparissias. Phytochemistry 1998, 48, 725–728. [Google Scholar] [CrossRef]
- Jiang, Y.; Tu, P.F. Xanthone O-glycosides from Polygala tenuifolia. Phytochemistry 2002, 60, 813–816. [Google Scholar] [CrossRef]
- Dao, T.T.; Dang, T.T.; Nguyen, P.H.; Kim, E.; Thuong, P.T.; Oh, W.K. Xanthones from Polygala karensium inhibit neuraminidases from influenza A viruses. Boorg. Med. Chem. Lett. 2012, 22, 3688–3692. [Google Scholar] [CrossRef] [PubMed]
- Dall’Acquaa, S.; Viola, G.; Cappelletti, E.M.; Innocenti, G. Xanthones from Polygala alpestris (Rchb.). Z. Für Nat. C 2004, 59, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Zheng, C.; Liu, Y.; Qu, L.; Yu, H.; Han, L.; Wang, T. Chemical and biological research on herbal medicines rich in xanthones. Molecules 2017, 22, 1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tizziani, T.; Venzke, D.; Ruani, A.P.; Pereira, M.; Micke, G.A.; Pizzolatti, M.G.; Brighente, I.M.C. Phytochemical and chemotaxonomic study of Polygala altomontana (Polygalaceae). Biochem. Syst. Ecol. 2018, 77, 1–3. [Google Scholar] [CrossRef]
- Tsujimoto, T.; Nishihara, M.; Osumi, Y.; Hakamatsuka, T.; Goda, Y.; Uchiyama, N.; Ozeki, Y. Structural analysis of polygalaxanthones, C-Glucosyl xanthones of Polygala tenuifolia roots. Chem. Pharm. Bull. 2019, 67, 1242–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Quang, T.; Dung, D.A.; Dong, N.T.; Hang, P.T.N.; Khoi, N.M.; Van Doan, V.; Nhiem, N.X. Sucrose esters and xanthones from Polygala karensium. Phytochem. Lett. 2020, 37, 75–79. [Google Scholar] [CrossRef]
- Tizziani, T.; Venzke, D.; Ruani, A.P.; Micke, G.A.; Pizzolatti, M.G.; Brighente, I.M.C. Dihydrostyryl-2-pyrone as a chemical marker of three non-xanthone-producing Polygala species (Polygalaceae). Biochem. Syst. Ecol. 2020, 90, 4034. [Google Scholar] [CrossRef]
- Zhang, D.; Miyase, T.; Kuroyanagı, M.; Umehara, K.; Ueno, A. Studies on the constituents of Polygala japonica Houtt. III. Structures of polygalasaponins XX-XXVII. Chem. Pharm. Bull. 1996, 44, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.L.; Zeng, K.W.; Shi, T.X.; Jiang, Y.; Tu, P.F. Sibiricasaponins A–E, five new triterpenoid saponins from the aerial parts of Polygala sibirica L. Fitoterapia 2013, 84, 295–301. [Google Scholar] [CrossRef]
- Jin, M.L.; Lee, D.Y.; Um, Y.; Lee, J.H.; Park, C.G.; Jetter, R.; Kim, O.T. Isolation and characterization of an oxidosqualene cyclase gene encoding a β-amyrin synthase involved in Polygala tenuifolia Willd. saponin biosynthesis. Plant. Cell. Rep. 2014, 33, 511–519. [Google Scholar] [CrossRef]
- Vinh, L.B.; Heo, M.; Phong, N.V.; Ali, I.; Koh, Y.S.; Kim, Y.H.; Yang, S.Y. Bioactive compounds from Polygala tenuifolia and their inhibitory effects on lipopolysaccharide-stimulated pro-inflammatory cytokine production in bone marrow-derived dendritic cells. Plants 2020, 9, 1240. [Google Scholar] [CrossRef]
- Lee, D.S.; Choi, H.G.; Li, B.; Kim, K.S.; Kim, S.A.; Chon, S.K.; Kim, Y.C. Neuroprotective effect of the acid hydrolysis fraction of the roots of Polygala tenuifolia. J. Physiol. Pathol. Korean Med. 2011, 25, 628–634. [Google Scholar]
- Li, Z.; Liu, Y.; Wang, L.; Liu, X.; Chang, Q.; Guo, Z.; Fan, T.P. Memory-enhancing effects of the crude extract of Polygala tenuifolia on aged mice. Evid-Basel Compl. Alt. 2014, 2014, 392324. [Google Scholar]
- Miyase, T.; Iwata, Y.; Ueno, A. Tenuifolioses G-P. Oligosaccharide Multi-Esters from the Root of Polygala tenuifolia Willd. Chem. Pharm. Bull. 1992, 40, 2741–2748. [Google Scholar] [CrossRef] [Green Version]
- Miyase, T.; Iwata, Y.; Ueno, A. Tenuifolioses A-F, oligosaccharide multi-esters from the roots of Polygala silia Willd. Chem. Pharm. Bull. 1991, 39, 3082–3084. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.H.; Doan, H.T.; Vu, T.V.; Pham, Q.T.; Khoi, N.M.; Huu, T.N.; Thuong, P.T. Oligosaccharide and glucose esters from the roots of Polygala arillata. Nat. Prod. Res. 2020, 34, 2900–2906. [Google Scholar] [CrossRef]
- Saitoh, H.; Miyase, T.; Ueno, A.; Atarashi, K.; Saiki, Y.; Senegoses, J.O. Oligosaccharide multi-esters from the roots of Polygala senega L. Chem. Pharm. Bull. 1994, 42, 641–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ba, Y.Y.; Wang, M.; Zhang, K.F.; Chen, Q.; Wang, J.; Lv, H.; Jiang, Y.Y.; Shi, R. Intestinal absorption profile of three Polygala oligosaccharide esters in Polygalae Radix and the effects of other components in Polygalae Radix on their absorption. Hindawi Evid-Basel Compl. Alt. 2019, 2019, 1379531. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Tu, P.F. Tenuifoliose Q, a new oligosaccharide ester from the root of Polygala tenuifolia Willd. J. Asian Nat. Prod. Res. 2003, 5, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, M.; Hostettmann, K. Hydroxycinnamic acid esters from Polygala chamaebuxus. Phytochemistry 1985, 24, 1793–1797. [Google Scholar] [CrossRef]
- Ardenghi, J.V.; Pretto, J.B.; Souza, M.M.; Junior, A.C.; Soldi, C.; Pizzolatti, M.G.; Santos, A.R. Antinociceptive properties of coumarins, steroid and dihydrostyryl-2-pyrones from Polygala sabulosa (Polygalaceae) in mice. J. Pharm. Pharmacol. 2006, 58, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.F.; Alves, C.Q.; Brandao, H.N.; David, J.M.; David, J.P.; Silva, R.L.; Oliveira, C.M. Poligalen, a new coumarin from Polygala boliviensis, reduces the release of TNF and IL-6 independent of NF-kB downregulation. Fitoterapia 2016, 113, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Do, J.C.; Yu, Y.J.; Jung, K.Y.; Son, K.H. Flavonoids from the Leaves of Polygalga japonica. Korean J. Pharmacogn. 1992, 23, 9–13. [Google Scholar]
- Rao, M.S.; Rao, P.S.; Kumar, J.K.; Raman, N.V. A rare flavonol glycoside from Polygala chinensis. Biochem. Syst. Ecol. 2003, 31, 635–636. [Google Scholar] [CrossRef]
- Rao, M.S.; Raman., M.V. A novel flavonoid from Polygala chinensis. Biochem. Syst. Ecol. 2004, 32, 447–448. [Google Scholar] [CrossRef]
- Sakthidevi, G.; Mohan, V.R. Comparative in vitro free radical scavenging activity of Polygala javana DC., Polygala chinensis L. and Polygala rosmarinifolia Wight & Arn. (Polygalaceae). J. Chem. Pharm. Sci. 2012, 2, 294–298. [Google Scholar]
- Shi, T.; Li, Y.; Jiang, Y.; Tu, P. Isolation of flavonoids from the aerial parts of Polygala tenuifolia Willd. and their antioxidant activities. J. Chin. Pharm. Sci. 2013, 22, 36. [Google Scholar] [CrossRef]
- Arruda-Silva, F.; Nascimento, M.V.P.; Luz, A.B.; Venzke, D.; Queiroz, G.S.; Fröde, T.S.; Dalmarco, E.M. Polygala molluginifolia A. St.-Hil. and Moq. prevent inflammation in the mouse pleurisy model by inhibiting NF-κB activation. Int. Immunopharmacol. 2014, 19, 334–341. [Google Scholar] [CrossRef]
- Yu-Hong, Z.H.O.U.; Zhang, S.Y.; Qiang, G.U.O.; Xing-Yun, C.H.A.I.; Jiang, Y.; Peng-Fei, T.U. Chemical investigation of the roots of Polygala sibirica L. Chin. J. Nat. Med. 2014, 12, 225–228. [Google Scholar]
- Borges, F.R.; Silva, M.D.; Córdova, M.M.; Schambach, T.R.; Pizzolatti, M.G.; Santos, A.R. Anti-inflammatory action of hydroalcoholic extract, dichloromethane fraction and steroid α-spinasterol from Polygala sabulosa in LPS-induced peritonitis in mice. J. Ethnopharmacol. 2014, 151, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, S.; Kumarswamy, C.; Chauhan, R.B.S. Lactonic lignans of Polygala chinensis. Phytochemistry 1973, 2550–2551. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201302212955 (accessed on 23 December 2021). [CrossRef]
- Ghosal, S.; Chauhan, R.P.; Srivastava, R.S. Structure of chinensin: A new lignan lactone from Polygala chinensis. Phytochemistry 1974, 13, 2281–2284. [Google Scholar] [CrossRef]
- Shibnath, G.; Ghosal, S. Two new Aryl Naphthalide Lignans from Polygala chinensis. Phytochemistry 1974, 13, 1933–1936. [Google Scholar]
- Hoffmann, J.J.; Wiedhopf, R.M.; Cole, J.R. Cytotoxic and tumor inhibitory agent from Polygala macradenia Gray (Polygalaceae): 4′-demethyldeoxypodophyllotoxin. J. Pharmacol. Sci. 1977, 66, 586–587. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, C.; Marston, A.; Wolfender, J.L.; Mavi, S.; Rogers, C.; Hostettmann, K. Isolation of polyphenols from Polygala gazensis and liquid chromatography–mass spectrometry of related African Polygala species. Phytochem. Anal. 1997, 8, 32–36. [Google Scholar] [CrossRef]
- Dall’Acqua, S.G.; Innocenti, G.; Viola, A.; Piovan, R.; Tumiato, E.M. Cappelletti, Cytotoxic compounds from Polygala vulgaris. Chem. Pharm. Bull. 2002, 50, 1499–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacaille-Dubois, M.A.; Delaude, C.; Mitaine-Offer, A.C. A review on the phytopharmacological studies of the genus Polygala. J. Ethnopharmacol. 2020, 249, 112417. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Fu, J.; Yang, J.; Zhang, D.; Yuan, Y.; Chen, N. Three triterpenoid saponins from the roots of Polygala japonica Houtt. Fitoterapia 2012, 83, 1184–1190. [Google Scholar] [CrossRef]
- Park, C.H.; Choi, S.H.; Koo, J.W.; Seo, J.H.; Kim, H.S.; Jeong, S.J.; Suh, Y.H. Novel cognitive improving and neuroprotective activities of Polygala tenuifolia Willdenow extract, BT-11. J. Neurosci Res. 2002, 70, 484–492. [Google Scholar] [CrossRef]
- Liu, P.; Hu, Y.; Guo, D.H.; Wang, D.X.; Tu, H.H.; Ma, L.; Xie, T.T.; Kong, L.Y. Potential antidepressant properties of Radix Polygalae (Yuan Zhi). Phytomedicine 2010, 17, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ma, C.; Li, B.M.; Sun, C. Polygala japonica Houtt. reverses depression-like behavior and restores reduced hippocampal neurogenesis in chronic stress mice. Biomed. Pharmacother. 2018, 99, 986–996. [Google Scholar] [CrossRef] [PubMed]
- Estrada, A.; Katselis, G.S.; Laarveld, B.; Barl, B. Isolation and evaluation of immunological adjuvant activities of saponins from Polygala senega L. Comp. Immunol. Microbiol. Infect. Dis. 2000, 23, 27–43. [Google Scholar] [CrossRef]
- Georgios, S.K.; Estrada, A.; Gorecki, D.K.J.; Barl, B. Adjuvant activities of saponins from the root of Polygala senega L. Can. J. Physiol. Pharmacol. 2007, 85, 1184–1194. [Google Scholar]
- Nagajyothi, P.C.; Cha, S.J.; Yang, I.J.; Sreekanth, T.V.; Kim, K.J.; Shin, H.M. Antioxidant and anti-inflammatory activities of zinc oxide nanoparticles synthesized using Polygala tenuifolia root extract. J. Photochem. Photobiol. B 2015, 146, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Zhang, G.D.; Li, F.Y.; Wang, T.L.; Suo, T.C.; Wang, C.H.; Li, Z.; Zhu, Y. Chemical Constituents from the Roots of Polygala arillata and Their Anti-Inflammatory Activities. J. Chem. 2019, 2019, 8079619. [Google Scholar] [CrossRef]
- Johann, S.; Mendes, G.M.; Missau, F.C.; Resende, M.A.; Pizzolatti, M.G. Antifungal Activity of Five Species of Polygala. Braz. J. Microbiol. 2011, 42, 1065–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tizziani, T.; Venzke, V.; Ruani, A.P.; Marques, L.B.; Prazeres, P.H.D.M.; Souza-Fagundes, E.M.; Pizzolatti, M.G.; Brighente, I.M.C. Antitumor screening of crude extracts of ten medicinal plants of Polygala genus from Southern Brazil. J. Appl. Phar. Sci. 2017, 7, 79–83. [Google Scholar]
- Lapa, F.R.; Gadotti, V.M.; Missau, F.C.; Pizzolatti, M.G.; Marques, M.C.A.; Dafré, A.L. Antinociceptive properties of the hydroalcoholic extract and the flavonoid rutin obtained from Polygala paniculata L. in mice. Basic Clin. Pharmacol. Toxicol. 2009, 104, 306–315. [Google Scholar] [CrossRef]
- Oh, J.J.; Kim, S.J. Inhibitory Effect of the root of Polygala tenuifolia on Bradykinin and COX 2-Mediated Pain and Inflammatory Activity. Trop. J. Pharm. Res. 2013, 12, 755–759. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Jia, M.; Wu, J.G.; Zhang, H.; Sun, L.N.; Chen, W.S.; Rahman, K. Anxiolytic and sedative-hypnotic activities of Polygala saponins from Polygala tenuifolia in mice. Pharm. Biol. 2010, 48, 1–7. [Google Scholar] [CrossRef]
- Duarte, F.S.; Duzzionim, M.; Mendes, B.G.; Pizzolatti, M.G.; Lima, T.C.M. Participation of dihydrostyryl-2-pyrones and styryl-2-pyrones in the central effects of Polygala sabulosa (Polygalaceae), a folk medicine topical anesthetic. Pharmacol. Biochem. Behav. 2007, 86, 150–161. [Google Scholar] [CrossRef]
- She, G.; Ba, Y.; Liu, Y.; Lv, H.; Wang, W.; Shi, R. Absorbable phenylpropenoyl sucroses from Polygala tenuifolia. Molecules 2011, 16, 5507–5513. [Google Scholar] [CrossRef]
- Quang, T.H.; Yen, D.T.H.; Dung, D.T.; Trang, D.T.; Ngan, N.T.T.; Van Kiem, P.; Van Minh, C. Anti-inflammatory phenylpropanoid glycosides from the roots of Polygala aureocauda Dunn. Vietnam J. Chem. 2019, 57, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.Y.; Liu, M.; Wu, Y.L.; Guo, Y.Q.; Li, Y.S. Separation and identification of new sucrose esters from root of Polygala tenuifolia Willd. J. Shenyang Pharm. Univ. 2010, 10. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-SYYD201010006.htm (accessed on 23 December 2021).
- Ikeya, Y.; Sugama, K.; Okada, M.; Mitsuhashi, H. Four new phenolic glycosides from Polygala tenuifolia. Chem. Pharm. Bull. 1991, 39, 2600–2605. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Miyase, T.; Kuroyanagi, M.; Umehara, K.; Noguchi, H. Oligosaccharide polyesters from roots of Polygala glomerata. Phytochemistry 1998, 47, 45–52. [Google Scholar] [CrossRef]
- Miyase, T.; Noguchi, H.; Chen, X.M. Sucrose esters and xanthone C-glycosides from the roots of Polygala sibirica. J. Nat. Prod. 1999, 62, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Hossin, A.Y.; Inafuku, M.; Takara, K.; Nugara, R.N.; Oku, H. Syringin: A Phenylpropanoid Glycoside Compound in Cirsium brevicaule A. Gray. Root Modulates Adipogenesis. Molecules 2021, 26, 1531. [Google Scholar] [CrossRef] [PubMed]
- Ersöz, T.; Saraçoğlu, İ.; Kırmızıbekmez, H.; Yalçın, F.N.; Harput, Ü.; Dönmez, A.A.; Çalış, İ. Iridoid Phenylethanoid and Phenol Glycosides from Phlomis chimerae. Hacet. Univ. J. Fac. Pharm. 2001, 21, 23–33. [Google Scholar]
- Dickey, E.E. Liriodendrin, a new lignan diglucoside from the inner bark of yellow poplar (Liriodendron tulipifera L.). J. Org. Chem. 1958, 23, 179–184. [Google Scholar] [CrossRef]
- Jianfeng, W.; Sibao, C.; Shilin, C.; Pengfei, T.; Lijun, W.; Lina, Y. Isolation and identification of chemical constituents of Polygala hongkongensis. Zhongcaoyao 2007, 38, 985–987. [Google Scholar]
- Chaudhuri, R.K.; Sticher, O. New Iridoid Glucosides and a Lignan Diglucoside from Globularia alypum L. Helv. Chim. Acta 1981, 64, 3–15. [Google Scholar] [CrossRef]
- Özgen, U.; Kazaz, C.; Secen, H.; Çalış, İ.; Coşkun, M.; Houghton, P.J. A novel naphthoquinone glycoside from Rubia peregrina L. Türk J. Chem. 2009, 33, 561–568. [Google Scholar]
- Panda, P.; Appalashetti, M.; MA Judeh, Z. Phenylpropanoid sucrose esters: Plant-derived natural products as potential leads for new therapeutics. Curr. Med. Chem. 2011, 18, 3234–3251. [Google Scholar] [CrossRef] [PubMed]
- Quang, T.H.; Cong, P.T.; Yen, D.T.H.; Nhiem, N.X.; Tai, B.H.; Yen, P.H.; Ngan, N.T.T.; Kim, D.G.; Kim, Y.C.; Oh, H.; et al. Triterpenoid saponins and phenylpropanoid glycosides from the roots of Polygala japonica Houtt. with anti-inflammatory activity. Phytochem. Lett. 2018, 24, 60–66. [Google Scholar] [CrossRef]
- Wang, N.; Yao, X.; Ishii, R.; Kitanaka, S. Bioactive sucrose esters from Bidens parviflora. Phytochemistry 2003, 62, 741–746. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Klinke, A.; Nussbaum, C.; Kubala, L.; Friedrichs, K.; Rudolph, T.K.; Rudolph, V.; Paust, H.J.; Schröder, C.; Benten, D.; Lau, D.; et al. Myeloperoxidase attracts neutrophils by physical forces. Blood. 2011, 117, 1350–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinecke, J.W. Tyrosyl radical production by myeloperoxidase: A phagocyte pathway for lipid peroxidation and dityrosine cross-linking of proteins. Toxicology 2002, 177, 11–22. [Google Scholar] [CrossRef]
- Galijasevic, S.; Saed, G.M.; Diamond, M.P.; Abu-Soud, H.M. Myeloperoxidase up-regulates the catalytic activity of inducible nitric oxide synthase by preventing nitric oxide feedback inhibition. Proc. Natl. Acad. Sci. USA 2003, 100, 14766–14771. [Google Scholar] [CrossRef] [Green Version]
- Wurtz, N.R.; Viet, A.; Shaw, S.A.; Dilger, A.; Valente, M.N.; Khan, J.A.; Jusuf, S.; Narayanan, R.; Fernando, G.; Lo, F.; et al. Potent Triazolopyridine Myeloperoxidase Inhibitors. ACS Med. Chem. Lett. 2018, 9, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Orlando, B.J.; Malkowski, M.G. Crystal structure of rofecoxib bound to human cyclooxygenase-2. Acta Crystallogr. F Struct. Biol. Commun. 2016, 72, 772–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcin, E.D.; Arvai, A.S.; Rosenfeld, R.J.; Kroeger, M.D.; Crane, B.R.; Andersson, G.; Andrews, G.; Hamley, P.J.; Mallinder, P.R.; Nicholls, D.J.; et al. Anchored plasticity opens doors for selective inhibitor design in nitric oxide synthase. Nat. Chem. Biol. 2008, 4, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippert, T.; Rarey, M. Fast automated placement of polar hydrogen atoms in protein-ligand complexes. J. Cheminformaticts 2009, 1, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bietz, S.; Urbaczek, S.; Schulz, B.; Rarey, M. Protoss: A holistic approach to predict tautomers and protonation states in protein-ligand complexes. J. Cheminformaticts 2014, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schellhammer, I.; Rarey, M.; Trix, X. Structure-based molecule indexing for large-scale virtual screening in sublinear time. J. Comput. Aid. Mol. Des. 2007, 21, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Henzler, A.M.; Urbaczek, S.; Hilbig, M.; Rarey, M. An integrated approach to knowledge-driven structure-based virtual screening. J. Comput. Aid. Mol. Des. 2014, 28, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Flachsenberg, F.; Meyder, A.; Sommer, K.; Penner, P.; Rarey, M. A Consistent Scheme for Gradient-Based Optimization of Protein-Ligand Poses. J. Chem. Inf. Model. 2020, 60, 6502–6522. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | |
|---|---|---|---|---|---|---|
| H–Atom | δH, ppm | δH, ppm | δH, ppm | δH, ppm | δH, ppm | δH, ppm |
| 1 | 5.53 d (3.8) | 5.52 d (3.8) | 5.45 d (3.4) | 5.45 d (3.5) | 5.44 d (3.6) | 5.46 d (3.6) |
| 2 | 3.51 dd (3.8/9.7) | 3.51 dd (3.8/9.7) | 3.46 (3.4/9.8) | 3.48 dd (3.5/9.5) | 3.45 dd (3.6/9.5) | 3.49 dd (3.6/9.5) |
| 3 | 3.70 t (9.2) | 3.69 t (9.2) | 3.68 t (9.3) | 3.68 t | 3.67 t (9.5) | 3.70 t (9.5) |
| 4 | 3.34 t (9.3) | 3.34 t (9.3) | 3.43 t (9.3) | 3.43 t (9.5) | 3.42 t (9.5) | 3.45 t (9.5) |
| 5 | 4.29 gdd (9.3/7.3) | 4.28 gdd (9.3/7.3) | 3.92 m | 3.92 m | 3.92 m | 3.94 m |
| 6 | 4.23 dd (11.7/7.3) 4.69 gd (11.7) | 4.23 dd (11.7/7.3) 4.69 gd (11.7) | 3.84 † 3.78 dd (12.2/4.4) | 3.85/3.79 | 3.84 dd (12.0/2.1) 3.78 dd (12.0/4.6) | 3.86 † 3.81 † |
| 1′ | 3.65 d (12.2) | 3.65 d (12.2) | 3.68 d (12.3) | 3.64 d (12.1) | 3.66 d (12.2) | 3.69 d (12.2) |
| 3.61 d (12.2) | 3.60 d (12.2) | 3.61 d (12.3) | 3.60 d (12.1) | 3.60 d (12.2) | 3.62 d (12.2) | |
| 2′ | - | - | - | - | - | - |
| 3′ | 5.53 d (8.1) | 5.54 d (8.1) | 5.49 d (7.8) | 5.49 d (7.8) | 5.49 d (7.8) | 5.50 d (7.8) |
| 4′ | 4.52 t (8.1) | 4.52 t (8.1) | 4.40 t (7.8) | 4.40 t | 4.39 t (7.8) | 4.41 t (7.8) |
| 5′ | 4.00 ddd (3.1/8.1/10.0) | 4.00 ddd (3.1/8.1/10.0) | 3.97 m | 3.97 m | 3.95 m | 3.97 m |
| 6′ | 3.91 dd (12.1/6.9) 3.84 † | 3.90 dd (12.1/6.9) 3.84 † | 3.85 † | 3.85 † | 3.86 † | 3.86 † |
| Acyl→Glu-6(OH) | SA | SA | - | - | - | - |
| 2″ | 6.88 s | 6.84 s | - | - | - | - |
| 6″ | 6.88 s | 6.84 s | - | - | - | - |
| 7″ | 7.57 d (15.7) | 7.56 d (15.9) | - | - | - | - |
| 8″ | 6.44 d (15.7) | 6.44 d (15.9) | - | - | - | - |
| 3″&5″-OMe | 3.85 s | 3.82 s | - | - | - | |
| Acyl→Fru-3′(OH) | SA | TMC | MFA | SA | TMC | FA |
| 2‴ | 6.84 s | 6.90 s | 7.22 d (2.0) | 6.93 s | 6.96 s | 7.22 d (2.0) |
| 5‴ | - | - | 6.95 d (8.0) | - | - | 6.8 d (8.2) |
| 6″″ | 6.84 s | 6.90 s | 7.21 dd (8.0/2.0) | 6.93 s | 6.96 s | 7.14 dd (8.2/2.0) |
| 7‴ | 7.65 d (15.7) | 7.67 d (15.9) | 7.71 d (15.9) | 7.69 d (15.9) | 7.72 d (16.0) | 7.71 d (15.9) |
| 8‴ | 6.42 d (15.7) | 6.53 d (15.9) | 6.47 d (15.9) | 6.44 d (15.9) | 6.54 d (16.0) | 6.44 d (15.9) |
| 3‴&5‴-OMe | 3.82 s (6H) | 3.84 s (6H) | 3.85 s (3H) | 3.87 s (6H) | 3.88 s (6H) | 3.89 s (3H) |
| 4‴-OMe | - | 3.77 s (3H) | 3.86 s (3H) | - | 3.80 s (3H) | - |
| 1 | 2 | 3 | 4 | 5 | 6 | ||
|---|---|---|---|---|---|---|---|
| C/H | DEPT | δC, ppm | δC, ppm | δC, ppm | δC, ppm | δC, ppm | δC, ppm |
| Glu | |||||||
| 1 | CH | 92.59 | 92.61 | 93.19 | 93.18 | 93.23 | 93.20 |
| 2 | CH | 72.99 | 72.97 | 72.94 | 72.79 | 73.00 | 72.94 |
| 3 | CH | 75.00 | 75.01 | 74.86 | 74.87 | 74.93 | 74.42 |
| 4 | CH | 71.83 | 71.81 | 71.02 | 70.99 | 71.07 | 71.01 |
| 5 | CH | 72.41 | 72.42 | 74.42 | 74.42 | 74.51 | 74.87 |
| 6 | CH2 | 65.60 | 65.58 | 62.20 | 62.20 | 62.28 | 62.18 |
| Fru | |||||||
| 1′ | CH2 | 63.72 | 63.72 | 65.19 | 65.20 | 65.24 | 65.19 |
| 2′ | C | 104.78 | 104.76 | 104.69 | 104.70 | 104.71 | 104.71 |
| 3′ | CH | 79.19 | 79.32 | 79.59 | 79.52 | 79.70 | 79.54 |
| 4′ | CH | 74.07 | 74.09 | 73.74 | 73.76 | 73.80 | 73.75 |
| 5′ | CH | 84.18 | 84.21 | 83.96 | 83.94 | 84.06 | 83.96 |
| 6′ | CH2 | 65.60 | 65.58 | 62.84 | 62.82 | 62.85 | 62.84 |
| Acyl→Glu-6(OH) | SA | SA | |||||
| 1″ | C | 126.46 | 126.48 | ||||
| 2″ | CH | 106.89 | 106.75 | ||||
| 3″ | C | 149.25 | 149.24 | ||||
| 4″ | C | 139.42 | 139.33 | ||||
| 5″ | C | 149.25 | 149.24 | ||||
| 6″ | CH | 106.89 | 106.75 | ||||
| 7″ | CH | 147.33 | 147.23 | ||||
| 8″ | CH | 115.65 | 115.66 | ||||
| 9″ | C | 169.16 | 169.10 | ||||
| 3″&5″-OMe | CH3 | 56.79 | 56.73 | ||||
| Acyl→Fru-3′(OH) | SA | TMC | MFA | SA | TMC | FA | |
| 1‴ | C | 126.51 | 131.34 | 128.56 | 126.49 | 131.46 | 127.60 |
| 2‴ | CH | 106.74 | 106.78 | 111.63 | 106.95 | 106.85 | 112.00 |
| 3″ | C | 149.24 | 154.59 | 150.47 | 149.27 | 154.66 | 150.49 |
| 4″″ | C | 139.30 | 141.10 | 152.68 | 139.43 | 141.17 | 149.21 |
| 5‴ | C/CH | 149.24 | 154.59 | 112.45 | 149.27 | 154.66 | 116.43 |
| 6‴ | CH | 106.74 | 106.78 | 124.10 | 106.95 | 106.85 | 124.15 |
| 7‴ | CH | 147.97 | 147.29 | 147.30 | 147.95 | 147.22 | 147.73 |
| 8‴ | CH | 115.29 | 117.63 | 115.83 | 115.32 | 117.74 | 114.89 |
| 9‴ | C | 168.31 | 167.85 | 168.17 | 168.26 | 167.82 | 168.39 |
| 3‴&5‴-OMe | CH3 | 56.84 | 56.79 | 56.51 | 56.90 | 56.79 | 56.88 |
| 4‴-OMe | CH3 | 61.18 | 61.39 | 61.20 |
| Protein | Ligand | JAMDA Score | All-Atom RMSD |
|---|---|---|---|
| MPO (PDB ID: 5QJ2) | PDB chemical ID: JXS | −2.32663 | 0.632 Å |
| Compound 1 | −3.78280 | ||
| Compound 2 | −3.57348 | ||
| Compound 3 | −2.90840 | ||
| Compound 4 | −3.13930 | ||
| Compound 5 | −2.97990 | ||
| Compound 6 | −2.83140 | ||
| Compound 7 | −2.50225 | ||
| Compound 8 | −3.03640 | ||
| Compound 9 | −3.52060 | ||
| iNOS (PDB ID: 3E7G) | PDB chemical ID: AT2 | −2.62362 | 0.682 Å |
| Compound 1 | −2.85444 | ||
| Compound 2 | −3.14240 | ||
| Compound 3 | −2.66980 | ||
| Compound 4 | −2.34210 | ||
| Compound 5 | −2.54230 | ||
| Compound 6 | −3.02600 | ||
| Compound 7 | −2.67380 | ||
| Compound 8 | −3.47560 | ||
| Compound 9 | −2.78550 | ||
| COX-2 (PDB ID: 5KIR) | PDB chemical ID: RCX | −2.52030 | 0.532 Å |
| Compound 1 | Positive | ||
| Compound 2 | Positive | ||
| Compound 3 | Positive | ||
| Compound 4 | Positive | ||
| Compound 5 | Positive | ||
| Compound 6 | Positive | ||
| Compound 7 | −2.56960 | ||
| Compound 8 | Positive | ||
| Compound 9 | Positive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ünlü, A.; Teralı, K.; Uğurlu Aydın, Z.; Dönmez, A.A.; Yusufoğlu, H.S.; Çalış, İ. Isolation, Characterization and In Silico Studies of Secondary Metabolites from the Whole Plant of Polygala inexpectata Peşmen & Erik. Molecules 2022, 27, 684. https://doi.org/10.3390/molecules27030684
Ünlü A, Teralı K, Uğurlu Aydın Z, Dönmez AA, Yusufoğlu HS, Çalış İ. Isolation, Characterization and In Silico Studies of Secondary Metabolites from the Whole Plant of Polygala inexpectata Peşmen & Erik. Molecules. 2022; 27(3):684. https://doi.org/10.3390/molecules27030684
Chicago/Turabian StyleÜnlü, Ayşe, Kerem Teralı, Zübeyde Uğurlu Aydın, Ali A. Dönmez, Hasan Soliman Yusufoğlu, and İhsan Çalış. 2022. "Isolation, Characterization and In Silico Studies of Secondary Metabolites from the Whole Plant of Polygala inexpectata Peşmen & Erik" Molecules 27, no. 3: 684. https://doi.org/10.3390/molecules27030684