
Characterization of Mitochondrial Prohibitin in Opsariichthys bidens and Its Potential Functions in Spermatogenesis

Abstract
:1. Introduction
2. Results
2.1. Histological Observations of Testis Development and Spermatogenesis in O. bidens
2.2. Full-Length cDNA Cloning and Protein Structure Prediction
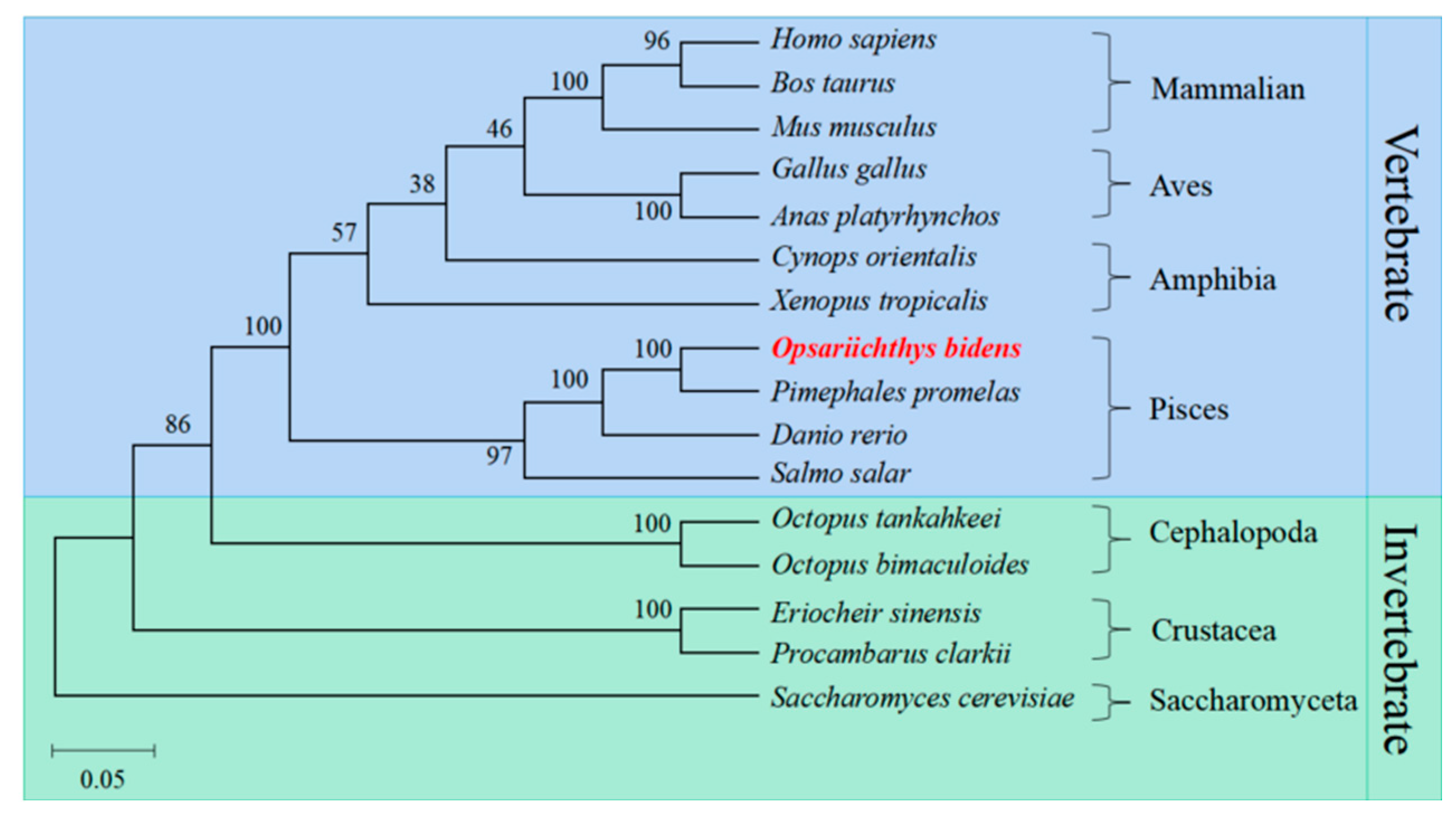
2.3. Multiple Sequence Alignment and Phylogenetic Analysis
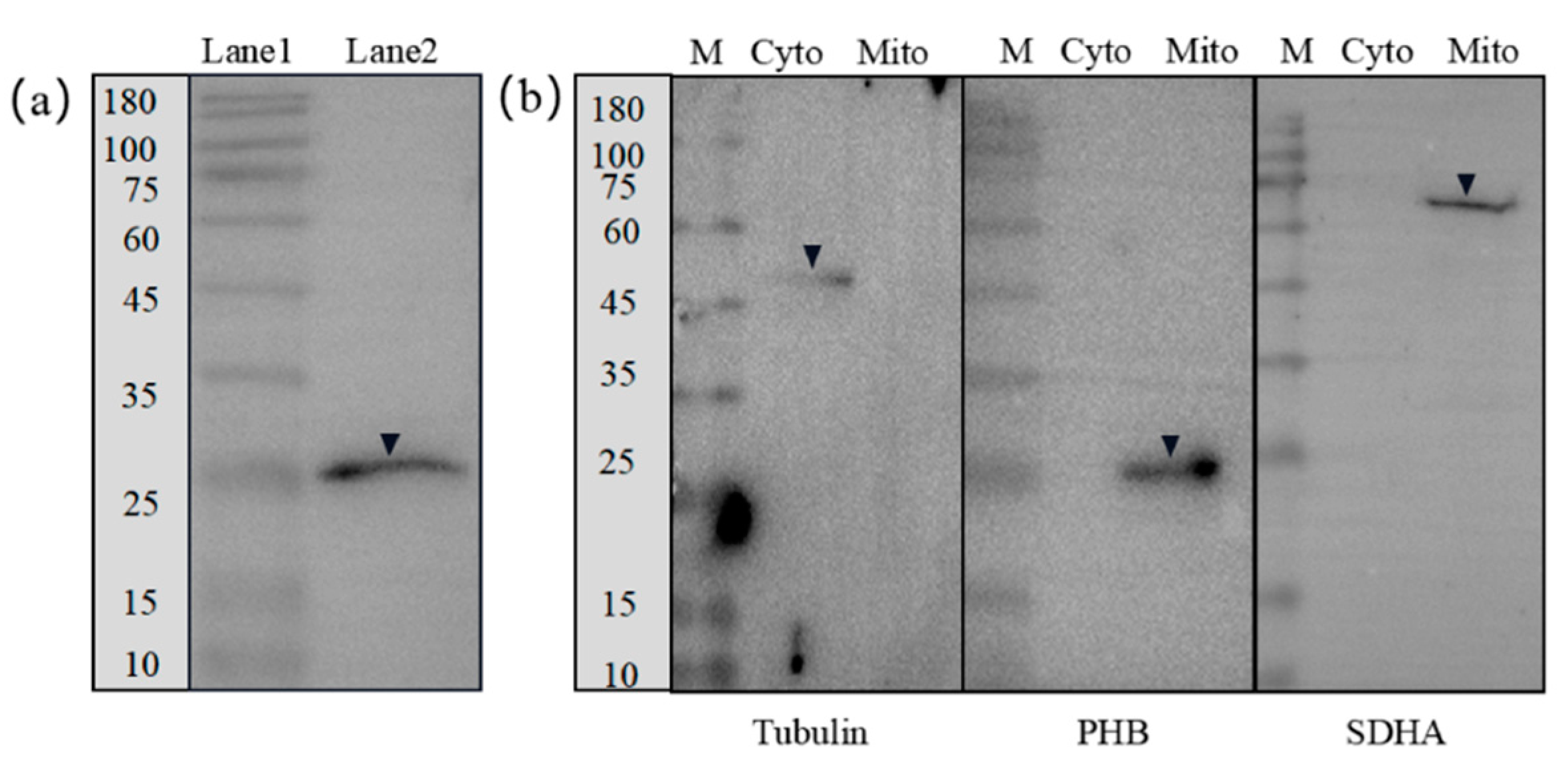
2.4. Expression Patterns of phb mRNA and PHB Protein
2.5. Subcellular Localization and Distribution of the Ob-PHB Protein during Spermatogenesis
3. Discussion
3.1. Testis Development and Spermatogenesis in O. bidens
3.2. High Conservation of PHB Protein
3.3. PHB Is Widely Expressed in Organisms and Participates in Spermatogenesis
3.4. PHB May Influence Mitochondrial Structure and Function and Mediate Mitochondrial Degradation during Spermatogenesis in O. bidens
4. Materials and Methods
4.1. Animal Preparation
4.2. Histological Analysis
4.3. Full-Length cDNA Cloning of Ob-phb
4.4. Analysis and Prediction of PHB Protein
4.5. Real-Time Quantitative PCR
4.6. Western Blotting
4.7. Extraction of Mitochondria and Detection of the PHB Protein
4.8. Immunofluorescence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Uribe, M.C.; Grier, H.J.; Mejía-Roa, V. Comparative testicular structure and spermatogenesis in bony fishes. Spermatogenesis 2015, 4, e983400. [Google Scholar] [CrossRef] [PubMed]
- Vertika, S.; Singh, K.K.; Rajender, S. Mitochondria, spermatogenesis, and male infertility—An update. Mitochondrion 2020, 54, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Cui, Y.; Guo, X.; Wang, L.; Li, S.; Lu, Y.; Bi, Y.; Huang, X.; Lin, M.; Xia, Y.; et al. 2,2’,4,4’-Tetrabromodiphenyl ether disrupts spermatogenesis, impairs mitochondrial function and induces apoptosis of early leptotene spermatocytes in rats. Reprod. Toxicol. 2015, 51, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Quan, C.; Shi, Y.; Wang, C.; Wang, C.; Yang, K. p,p’-DDE damages spermatogenesis via phospholipid hydroperoxide glutathione peroxidase depletion and mitochondria apoptosis pathway. Environ. Toxicol. 2016, 31, 593–600. [Google Scholar] [CrossRef] [PubMed]
- De Lamirande, E.; Cagnon, C. Human sperm hyperactivation and capacitation as parts of an oxidative process. Free Radic. Biol. Med. 1993, 14, 157–166. [Google Scholar] [CrossRef]
- Alvarez, J.G.; Storey, B.T. Differential incorporation of fatty acids into and peroxidative loss of fatty acids from phospholipids of human spermatozoa. Mol. Reprod. Dev. 1995, 42, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Ankel-Simons, F.; Cummins, J.M. Misconceptions about mitochondria and mammalian fertilization: Implications for theories on human evolution. Proc. Natl. Acad. Sci. USA 1996, 93, 13859–13863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummins, J. Mitochondrial DNA in mammalian reproduction. Rev. Reprod. 1998, 3, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fabiani, M.B.; Montanaro, M.A.; Stringa, P.; Lacunza, E.; Cattaneo, E.R.; Santana, M.; Pellon-Maison, M.; Gonzalez-Baro, M.R. Glycerol-3-phosphate acyltransferase 2 is essential for normal spermatogenesis. Biochem. J. 2017, 474, 3093–3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puglisi, R.; Tramer, F.; Carlomagno, G.; Gandini, L.; Panfili, E.; Stefanini, M.; Lenzi, A.; Mangia, F.; Boitani, C. PHGPx in spermatogenesis: How many functions? Contraception 2005, 72, 291–293. [Google Scholar] [CrossRef]
- Gu, N.-H.; Zhao, W.-L.; Wang, G.-S.; Sun, F. Comparative analysis of mammalian sperm ultrastructure reveals relationships between sperm morphology, mitochondrial functions and motility. Reprod. Biol. Endocrinol. 2019, 17, 66. [Google Scholar] [CrossRef] [Green Version]
- Ferramosca, A.; Provenzano, S.P.; Montagna, D.D.; Coppola, L.; Zara, V. Oxidative stress negatively affects human sperm mitochondrial respiration. Urology 2013, 82, 78–83. [Google Scholar] [CrossRef]
- Ulloa-Rodríguez, P.; Figueroa, E.; Díaz, R.; Lee-Estevez, M.; Short, S.; Farías, J.G. Mitochondria in teleost spermatozoa. Mitochondrion 2017, 34, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Kihara, A.; Akiyama, Y.; Ito, K.A. protease complex in the Escherichia coli plasma membrane: HflKC (HflA) forms a complex with FtsH (HflB), regulating its proteolytic activity against SecY. EMBO J. 1996, 15, 6122–6131. [Google Scholar] [CrossRef]
- McClung, J.K.; Jupe, E.R.; Liu, X.T.; dell’Orco, R.T. Prohibitin: Potential role in senescence, development, and tumor suppression. Exp. Gerontol. 1995, 30, 99–124. [Google Scholar] [CrossRef]
- Snedden, W.A.; Fromm, H. Characterization of the plant homologue of prohibitin, a gene associated with antiproliferative activity in mammalian cells. Plant Mol. Biol. 1997, 3, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Merkwirth, C.; Langer, T. Prohibitin function within mitochondria: Essential roles for cell proliferation and cristae morphogenesis. Biochim. Biophys. Acta. 2009, 1793, 27–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijtmans, L.G.; Artal, S.M.; Grivell, L.A.; Coates, P.J. The mitochondrial PHB complex: Roles in mitochondrial respiratory complex assembly, ageing and degenerative disease. Cell. Mol. Life Sci. 2002, 59, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, T.; Model, K.; Langer, T. Formation of membrane-bound ring complexes by prohibitins in mitochondria. Mol. Biol. Cell. 2005, 16, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Artal-Sanz, M.; Tsang, W.Y.; Willems, E.M.; Grivell, L.A.; Lemire, B.D.; van der Spek, H.; Nijtmans, L.G. The mitochondrial prohibitin complex is essential for embryonic viability and germline function in Caenorhabditis elegans. J. Biol. Chem. 2003, 278, 32091–32099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coates, P.J.; Jamieson, D.J.; Smart, K.; Prescott, A.R.; Hall, P.A. The prohibitin family of mitochondrial proteins regulate replicative lifespan. Curr. Biol. 1997, 7, 607–610. [Google Scholar] [CrossRef] [Green Version]
- Berger, K.H.; Yaffe, M.P. Prohibitin family members interact genetically with mitochondrial inheritance components in Saccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 4043–4052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eveleth, D.D., Jr.; Marsh, J.L. Sequence and expression of the Cc gene, a member of the dopa decarboxylase gene cluster of Drosophila: Possible translational regulation. Nucleic Acids Res. 1986, 14, 6169–6183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijtmans, L.G.; de Jong, L.; Artal Sanz, M.; Coates, P.J.; Berden, J.A.; Back, J.W.; Muijsers, A.O.; van der Spek, H.; Grivell, L.A. Prohibitins act as a membrane-bound chaperone for the stabilization of mitochondrial proteins. EMBO J. 2000, 19, 2444–2451. [Google Scholar] [CrossRef] [Green Version]
- Kasashima, K.; Sumitani, M.; Satoh, M.; Endo, H. Human prohibitin 1 maintains the organization and stability of the mitochondrial nucleoids. Exp. Cell. Res. 2008, 314, 988–996. [Google Scholar] [CrossRef]
- Steglich, G.; Neupert, W.; Langer, T. Prohibitins regulate membrane protein degradation by the m-AAA protease in mitochondria. Mol. Cell. Biol. 1999, 19, 3435–3442. [Google Scholar] [CrossRef] [Green Version]
- Merkwirth, C.; Dargazanli, S.; Tatsuta, T.; Geimer, S.; Löwer, B.; Wunderlich, F.T.; von Kleist-Retzow, J.C.; Waisman, A.; Westermann, B.; Langer, T. Prohibitins control cell proliferation and apoptosis by regulating OPA1-dependent cristae morphogenesis in mitochondria. Genes. Dev. 2008, 22, 476–488. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.; Zhang, H.; Huang, B.; Liu, Q.; Wang, Y.; Shi, D.; Li, X. Expression pattern of prohibitin, capping actin protein of muscle Z-line beta subunit and tektin-2 gene in Murrah buffalo sperm and its relationship with sperm motility. Asian-Australas. J. Anim. Sci. 2018, 31, 1729–1737. [Google Scholar] [CrossRef] [Green Version]
- Hernando-Rodríguez, B.; Artal-Sanz, M. Mitochondrial Quality Control Mechanisms and the PHB (Prohibitin) Complex. Cells 2018, 7, 238. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.L.; Hou, C.-C.; Yang, W.-X. Mitochondrial prohibitin and its ubiquitination during crayfish Procambarus clarkii spermiogenesis. Cell. Tissue. Res. 2015, 359, 679–692. [Google Scholar] [CrossRef]
- Mao, H.; Wang, D.-H.; Zhou, H.; Yang, W.-X. Characterization and expression analysis of prohibitin in the testis of Chinese mitten crab Eriocheir sinensis. Mol. Biol. Rep. 2012, 39, 7031–7039. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-R.; Fan, Y.-S.; Yang, W.-X. Mitochondrial prohibitin and its ubiquitination during spermatogenesis of the swimming crab Charybdis japonica. Gene 2017, 627, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Du, C.; Zheng, X.; Hou, C.; Wang, Y.; Xu, S.; Yang, Y.; Zhu, J.; Jin, S. Characterisation, expression and possible functions of prohibitin during spermatogenesis in the silver pomfret Pampus argenteus. Reprod. Fertil. Dev. 2020, 32, 1084–1098. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhao, Y.-Q.; Han, Y.-L.; Hou, C.-C.; Zhu, J.-Q. Characterization of mitochondrial prohibitin from Boleophthalmus pectinirostris and evaluation of its possible role in spermatogenesis. Fish. Physiol. Biochem. 2017, 43, 1299–1313. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Gao, X.; Lin, C.; Feng, B.; Hou, C.; Zhu, J.; Wang, J. Cytological features of spermatogenesis in Opsariichthys bidens (Teleostei, Cyprinidae). Anim. Reprod. Sci. 2020, 222, 106608. [Google Scholar] [CrossRef] [PubMed]
- Huszno, J.; Klag, J. The reproductive cycle in the male gonads of Danio rerio (Teleostei, Cyprinidae). Stereological analysis. Micron. 2012, 43, 666–672. [Google Scholar] [CrossRef]
- Xie, N.Y.; Yang, Q.F.; He, X.F. The embryonic and larval development of Tor brevififilis. J. Fisheries China 2002, 26, 115–121. [Google Scholar]
- Fu, S.-Y.; Jiang, J.-H.; Yang, W.-X.; Zhu, J.-Q. A histological study of testis development and ultrastructural features of spermatogenesis in cultured Acrossocheilus fasciatus. Tissue Cell 2016, 48, 49–62. [Google Scholar] [CrossRef]
- Mattei, X.; Slau, Y.; Thiaw, O.T.; Thiam, D. Peculiarities in the organization of testis of Ophidion sp. (Pisces: Teleostei). Evidence for two types of spermatogenesis in teleost fish. J. Fish Biol. 1993, 43, 931–937. [Google Scholar] [CrossRef]
- Rupik, W.; Huszno, J.; Klag, J. Cellular organisation of the mature testes and stages of spermiogenesis in Danio rerio (Cyprinidae; Teleostei)—structural and ultrastructural studies. Micron 2011, 42, 833–839. [Google Scholar] [CrossRef]
- Andrade, R.F.; Bazzoli, N.; Rizzo, E.; Sato, Y. Continuous gametogenesis in the neotropical freshwater teleost, Bryconops affinis (Pisces:Characidae). Tissue Cell 2001, 33, 524–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalho-Santos, J.; Varum, S.; Amaral, S.; Mota, P.C.; Sousa, A.P.; Amaral, A. Mitochondrial functionality in reproduction: From gonads and gametes to embryos and embryonic stem cells. Hum. Reprod Update 2009, 15, 553–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rato, L.; Alves, M.G.; Socorro, S.; Duarte, A.I.; Cavaco, J.E.; Oliveira, P.F. Metabolic regulation is important for spermatogenesis. Nat. Rev. Urol. 2012, 9, 330–338. [Google Scholar] [CrossRef]
- Varuzhanyan, G.; Rojansky, R.; Sweredoski, M.J.; Graham, R.L.; Hess, S.; Ladinsky, M.S.; Chan, D.C. Mitochondrial fusion is required for spermatogonial differentiation and meiosis. eLife 2019, 8, e51601. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaka, T.; Kosako, H.; Yoshizumi, T.; Furukawa, R.; Hirano, Y.; Kuge, O.; Tamada, T.; Koshiba, T. Structural Basis of Mitochondrial Scaffolds by Prohibitin Complexes: Insight into a Role of the Coiled-Coil Region. iScience 2019, 19, 1065–1078. [Google Scholar] [CrossRef]
- Hou, C.-C.; Wei, C.-G.; Lu, C.-P.; Gao, X.-M.; Yang, W.-X.; Zhu, J.-Q. Prohibitin-mediated mitochondrial ubiquitination during spermiogenesis in Chinese mitten crab Eriocheir sinensis. Oncotarget 2017, 8, 98782–98797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.-T.; Wang, D.-H.; Lan, Z.; Zhou, H.; Yang, W.-X. Gene expression profiles of prohibitin in testes of Octopus tankahkeei (ot-phb) revealing its possible role during spermiogenesis. Mol. Biol. Rep. 2012, 39, 5519–5552. [Google Scholar] [CrossRef]
- Ikonen, E.; Fiedler, K.; Parton, R.G.; Simons, K. Prohibitin, an antiproliferative protein, is localized to mitochondria. FEBS Lett. 1995, 358, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Kasashima, K.; Ohta, E.; Kagawa, Y.; Endo, H. Mitochondrial functions and estrogen receptor-dependent nuclear translocation of pleiotropic human prohibitin 2. J. Biol Chem. 2006, 281, 36401–36410. [Google Scholar] [CrossRef] [Green Version]
- Shibanuma, M.; Inoue, A.; Ushida, K.; Uchida, T.; Ishikawa, F.; Mori, K.; Nose, K. Importance of mitochondrial dysfunction in oxidative stress response: A comparative study of gene expression profiles. Free Radic. Res. 2011, 45, 672–680. [Google Scholar] [CrossRef]
- Wang, M.-J.; Ou, J.-X.; Chen, G.-W.; Wu, J.-P.; Shi, H.-J.; O, W.S.; Martin-DeLeon, P.A.; Chen, H. Does prohibitin expression regulate sperm mitochondrial membrane potential, sperm motility, and male fertility? Antioxid. Redox Signal. 2012, 17, 513–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, R.R.; Chen, G.W.; Shi, H.J.; O, W.S.; Martin-DeLeon, P.A.; Chen, H. Prohibitin involvement in the generation of mitochondrial superoxide at complex I in human sperm. J. Cell. Mol. Med. 2017, 21, 121–129. [Google Scholar] [CrossRef] [PubMed]
- McClung, J.K.; Danner, D.B.; Stewart, D.A.; Smith, J.R.; Schneider, E.L.; Lumpkin, C.K.; dell’Orco, R.T.; Nuell, M.J. Isolation of a cDNA that hybrid selects antiproliferative mRNA from rat liver. Biochem. Biophys. Res. Commun. 1989, 164, 1316–1322. [Google Scholar] [CrossRef]
- Kang, T.; Lu, W.; Xu, W.; Anderson, L.; Bacanamwo, M.; Thompson, W.; Chen, Y.E.; Liu, D. MicroRNA-27 (miR-27) targets prohibitin and impairs adipocyte differentiation and mitochondrial function in human adipose-derived stem cells. J. Biol. Chem. 2013, 288, 34394–34402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixit, V.D.; Sridaran, R.; Edmonsond, M.A.; Taub, D.; Thompson, W.E. Gonadotropin-releasing hormone attenuates pregnancy-associated thymic involution and modulates the expression of antiproliferative gene product prohibitin. Endocrinology 2003, 144, 1496–1505. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.D.; Gao, X.M.; Zhao, Y.Q.; Hou, C.C.; Zhu, J.Q. The C-terminal kinesin motor KIFC1 may participate in nuclear reshaping and flagellum formation during spermiogenesis of Larimichthys crocea. Fish Physiol. Biochem. 2017, 43, 1351–1371. [Google Scholar] [CrossRef]
- Lin, C.; Tang, D.; Gao, X.; Jiang, H.; Du, C.; Zhu, J. Molecular characterization, dynamic transcription, and potential function of KIF3A/KIF3B during spermiogenesis in Opsariichthys bidens. Gene 2021, 789, 145795. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer/Probe | Sequence (5′-3′) | Purpose |
|---|---|---|
| PHBF1 | TTTGTGGTAGAAAAGGCAG | PCR |
| PHBF2 | AGTTTTCTTGCTCTCTTCCT | PCR |
| PHBR1 | GTGGTAATTGAAGGAGCG | PCR |
| PHBR2 | ATTAGCCACTTGCGGAT | PCR |
| 3′PHBF1 | CTCGCAATGTTACCTACCTCCC | 3′ RACE |
| 3′PHBF2 | GGAAAAGATGATGTCTGTAGAACCC | 3′ RACE |
| 3′PHBF3 | TCTTGCTCTCTTCCTGAAAGTATGG | 3′ RACE |
| 5′PHBR1 | GGTAGGTAACATTGCGAGAGCGG | 5′ RACE |
| 5′PHBR2 | CCTGCTTCTGCTGTTCTGCCTTTT | 5′ RACE |
| 5′PHBR3 | AGATGGGAGGTAGGTAACATTGCGA | 5′ RACE |
| qPCR-F | GAAGCCAATCATCTTTGACTGC | qPCR |
| qPCR-R | TCGGAACAGGATTCTTAGCGT | qPCR |
| β-actin F | TCCGTGACATCAAGGAGAAGC | qPCR |
| β-actin R | GGCAACGGAAACGCTCATT | qPCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Wang, J.; Gao, X.; Du, C.; Hou, C.; Zhang, C.; Zhu, J.; Tang, D. Characterization of Mitochondrial Prohibitin in Opsariichthys bidens and Its Potential Functions in Spermatogenesis. Int. J. Mol. Sci. 2022, 23, 7295. https://doi.org/10.3390/ijms23137295
Wang L, Wang J, Gao X, Du C, Hou C, Zhang C, Zhu J, Tang D. Characterization of Mitochondrial Prohibitin in Opsariichthys bidens and Its Potential Functions in Spermatogenesis. International Journal of Molecular Sciences. 2022; 23(13):7295. https://doi.org/10.3390/ijms23137295
Chicago/Turabian StyleWang, Li, Jingqian Wang, Xinming Gao, Chen Du, Congcong Hou, Chundan Zhang, Junquan Zhu, and Daojun Tang. 2022. "Characterization of Mitochondrial Prohibitin in Opsariichthys bidens and Its Potential Functions in Spermatogenesis" International Journal of Molecular Sciences 23, no. 13: 7295. https://doi.org/10.3390/ijms23137295