
Morphology, Palynology and Molecular Phylogeny of Barleria cristata L. (Acanthaceae) Morphotypes from India

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling and Morphological Analysis
2.2. Palynology
2.3. DNA Extraction, PCR and DNA Sequencing
2.4. Sequence Alignment and Phylogenetic Analyses
3. Results
3.1. Morphological Analyses
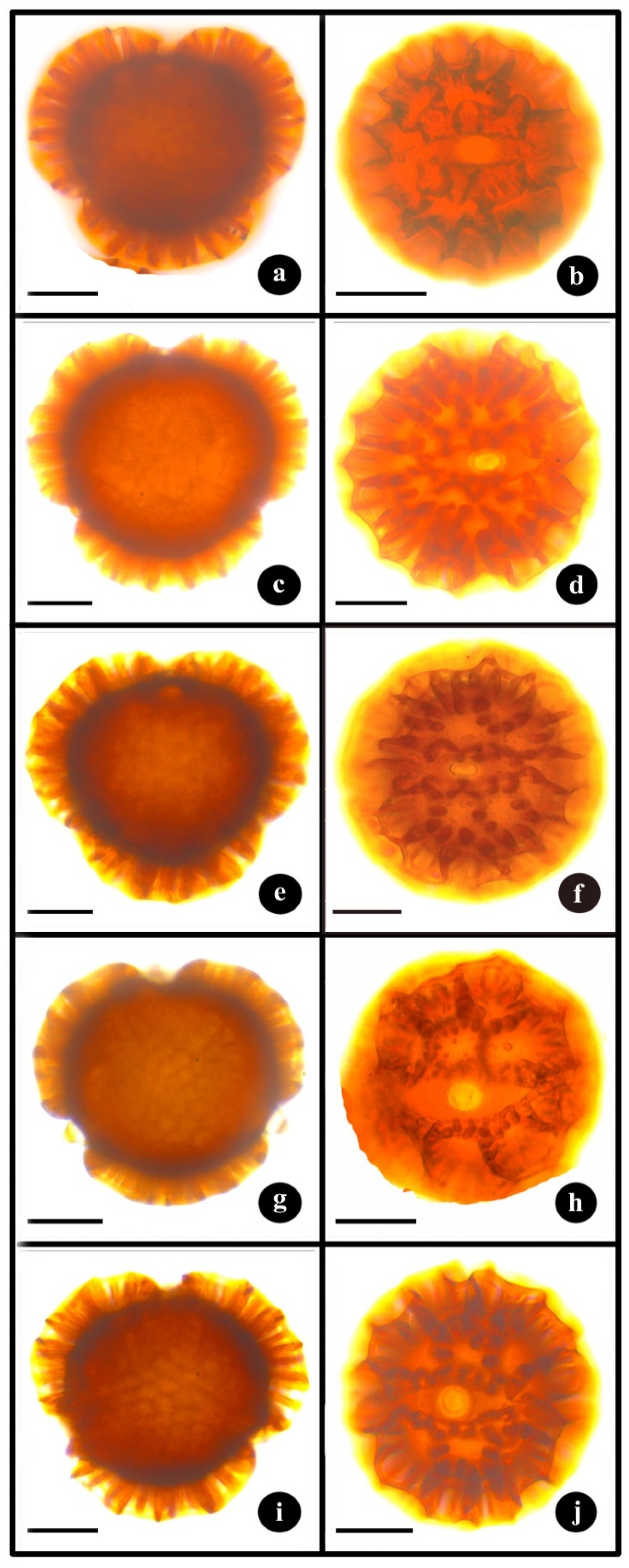
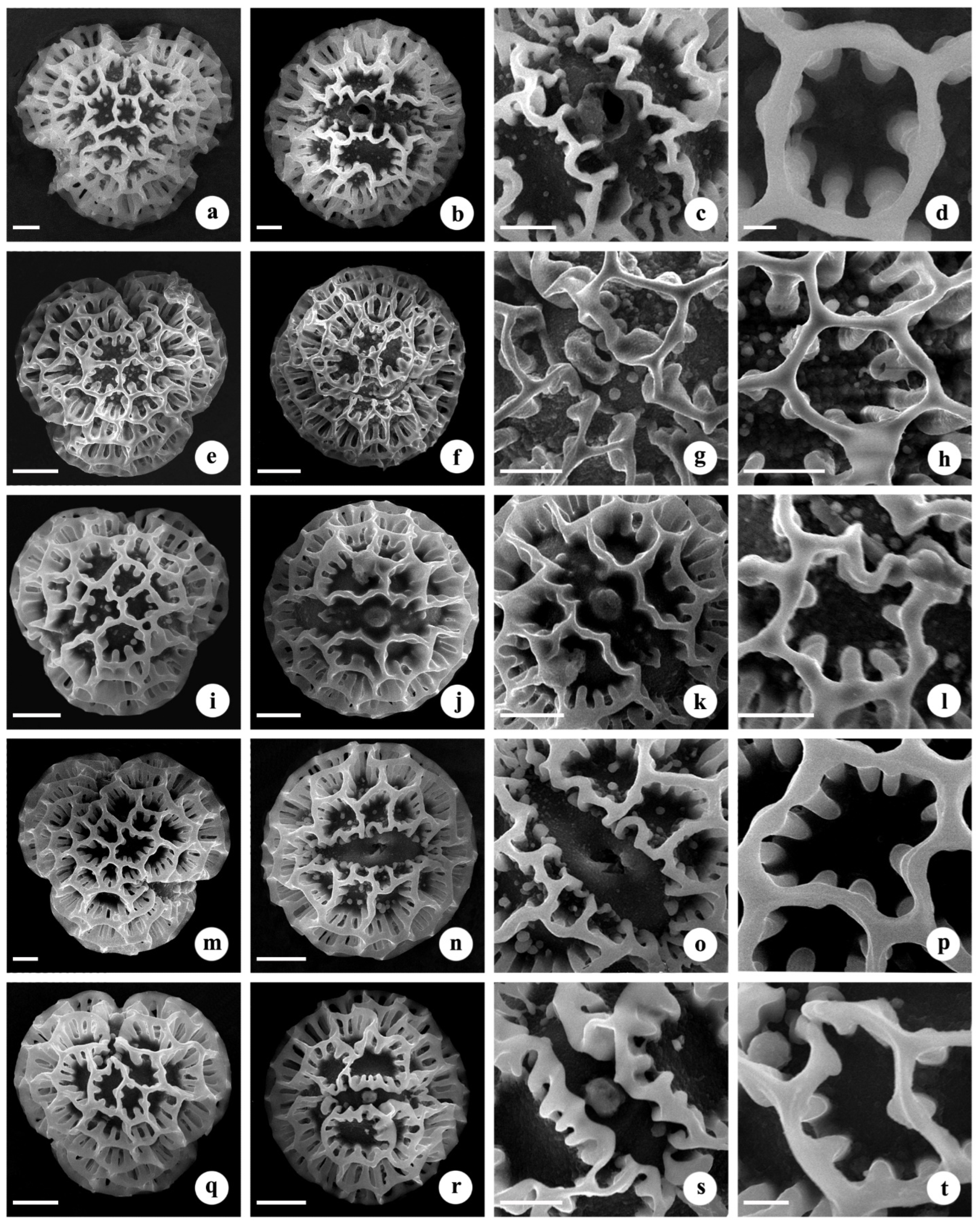
3.2. Palynology
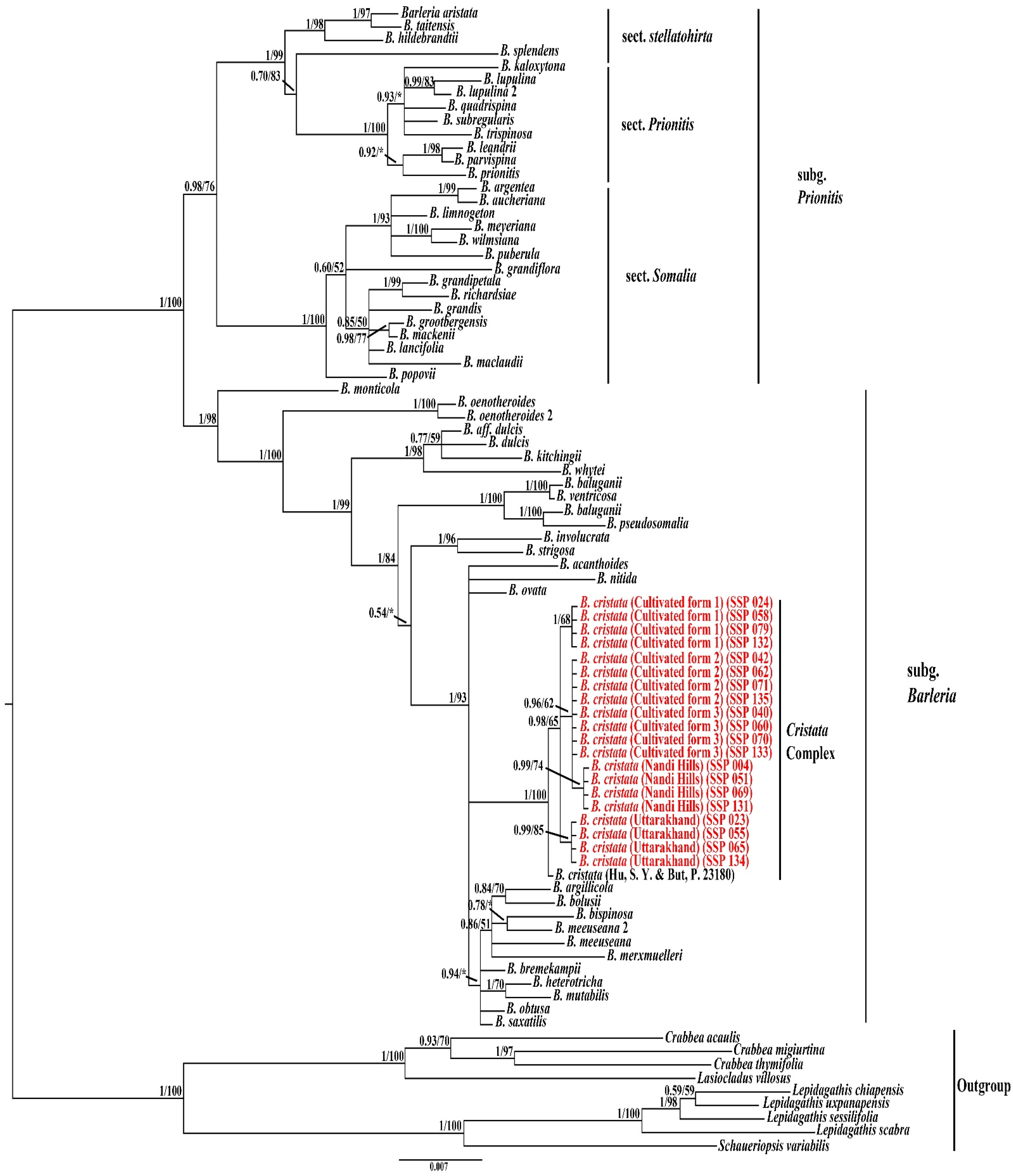
3.3. Phylogenetic Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darbyshire, I.; Fisher, A.E.; Kiel, C.A.; McDade, L.A. Phylogenetic relationships among species of Barleria (Acanthaceae, Lamiales): Molecular data reveal complex patterns of morphological evolution and support a revised classification. Taxon 2019, 68, 92–111. [Google Scholar] [CrossRef]
- Darbyshire, I.; Luke, Q. Barleria mirabilis (Acanthaceae): A remarkable new tree species from west Tanzania. Kew Bull. 2016, 71, 13. [Google Scholar] [CrossRef]
- Balkwill, M.J.; Balkwill, K. Delimitation and infra-generic classification of Barleria (Acanthaceae). Kew Bull. 1997, 52, 535–573. [Google Scholar] [CrossRef]
- Comito, R.; Darbyshire, I.; Kiel, C.; McDade, L.; Fisher, A.E. A RADseq phylogeny of Barleria (Acanthaceae) resolves fine-scale relationships. Mol. Phylogenet. Evol. 2022, 169, 107428. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.-H.; Liu, B.; Liang, Y.-S.; Ye, J.-F.; Saqib, S.; Meng, Z.; Lu, L.-M.; Chen, Z.-D. An updated Chinese vascular plant tree of life: Phylogenetic diversity hotspots revisited. J Syst. Evol. 2020, 58, 663–672. [Google Scholar] [CrossRef]
- Balkwill, M.J.; Balkwill, K. A preliminary analysis of distribution patterns in a large, pantropical genus, Barleria, L. (Acanthaceae). J. Biogeogr. 1998, 25, 95–110. [Google Scholar] [CrossRef]
- Shendage, S.M.; Yadav, S.R. Revision of the genus Barleria (Acanthaceae) in India. Rheedea 2010, 20, 81–130. [Google Scholar]
- Kumar, H.; Agrawal, R.; Kumar, V. Barleria cristata: Perspective towards phytopharmacological aspects. J. Pharm. Pharmacol. 2018, 70, 475–487. [Google Scholar] [CrossRef]
- Lekhak, M.M.; Patil, S.S.; Deshmukh, P.V.; Lekhak, U.M.; Vijay, K.; Anshu, R. Genus Barleria L. (Acanthaceae): A review of its taxonomy, cytogenetics, phytochemistry and pharmacological potential. J. Pharm. Pharmacol. 2022, 74, 812–842. [Google Scholar]
- Hickey, M.; King, C. The Cambridge Illustrated Glossary of Botanical Terms; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Ridgway, R. Color Standards and Color Nomenclature. Washington, DC, USA, 1912. 43p. Available online: https://library.si.edu/digital-library/book/colorstandardsc00ridg (accessed on 20 March 2022).
- Erdtman, G. The acetolysis method, a revised description. Sven. Bot. Tidskr. 1960, 54, 561–564. [Google Scholar]
- Yadav, P.B.; Manning, J.C.; Yadav, S.R.; Lekhak, M.M. A cytotaxonomic revision of Drimia Jacq. (Hyacinthaceae: Urgineoideae) in India. S. Afr. J. Bot. 2019, 123, 62–86. [Google Scholar] [CrossRef]
- Erdtman, G. Handbook of Palynology-An Introduction to the Study of Pollen Grains and Spores; Munksgaard: Copenhagen, Denmark, 1969. [Google Scholar]
- Erdtman, G. Pollen Morphology and Pollen Taxonomy; Hafner Publishing Company: New York, NY, USA, 1971. [Google Scholar]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Paterson, A.; Brubaker, C.; Wendel, J. A rapid method for extraction of cotton (Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis. Plant Mol. Biol. Rep. 1993, 11, 122–127. [Google Scholar] [CrossRef]
- Tamboli, A.S.; Dalavi, J.V.; Kadam, S.K.; Yadav, S.R.; Govindwar, S.P.; Simoes, A.R.G. New molecular phylogenetic evidence for Indian endemic species of the tribe Merremieae. Convolvulaceae. Plant Biosyst. 2021, 156, 440–449. [Google Scholar] [CrossRef]
- Hamilton, M.B. Four primer pairs for the amplification of chloroplast regions with intraspecific variation. Molec. Ecol. 1999, 8, 521–523. [Google Scholar]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose non-coding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Amer. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F.; Campbell, C.S.; Donoghue, M.J. The ITS region of nuclear ribosomal DNA: A valuable source of evidence on angiosperm phylogeny. Ann. Missouri. Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Sun, Y.; Skinner, D.Z.; Liang, G.H.; Hulbert, S.H. Phylogenetic analysis of Sorghum and related taxa using internal transcribed spacers of nuclear ribosomal DNA. Theoret. Appl. Genet. 1994, 89, 26–32. [Google Scholar] [CrossRef]
- McDade, L.A.; Daniel, T.F.; Kiel, C.A. Toward a comprehensive understanding of phylogenetic relationships among lineages of Acanthaceae s.l. (Lamiales). Am. J. Bot. 2008, 95, 1136–1152. [Google Scholar] [CrossRef]
- Gene Codes Corporation. Sequencher 5.1 Gene Codes Corporation. Ann Arbor, Michigan. 2012. Available online: http://genecodescom/ (accessed on 20 March 2022).
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinfo. 2004, 5, 113. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Bio. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multigene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat Methods 2012, 9, 772–775. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Devi, G.V.; Mathew, P.M. Cytological studies in the south Indian Acanthaceae: I. Genus Barleria L. Cytologia 1991, 56, 353–357. [Google Scholar] [CrossRef]
- Ranganath, R.M.; Krishnappa, D.G. Karyotypic studies in a few species of Barleria L. (Acanthaceae) from South India. Cytologia 1990, 55, 175–179. [Google Scholar] [CrossRef]
- Gavade, S.K.; Surveswaran, S.; Van-der-Maesen, L.J.G.; Lekhak, M.M. Taxonomic revision and molecular phylogeny of Flemingia subgenus Rhynchosioides (Leguminosae). Blumea 2019, 64, 253–271. [Google Scholar] [CrossRef]
- Al-Hakimi, A.S.; Faridah, Q.Z.; Abdulwahab, A.S.; Latiff, A. Pollen and seed morphology of Barleria L. (Barlerieae: Ruellioideae: Acanthaceae) of Yemen. S. Afr. J. Bot. 2018, 116, 185–191. [Google Scholar] [CrossRef]
- Nyirenda, F.C.; Balkwill, K. Significance of variation and evaluation of potential characters in three morphologically similar species of Barleria sect. Barleria (Acanthaceae) in southern Africa. Plant Syst. Evol. 2018, 304, 77–92. [Google Scholar] [CrossRef]
- Cornejo, A.M.B.; Lindqvist, C.; Balch, E.M.P.M.; Delgado, M.E.S. Phylogeny of the Stachys coccinea (Lamiaceae) Complex Based on Molecular and Morphological Data. Sys. Bot. 2017, 42, 484–493. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
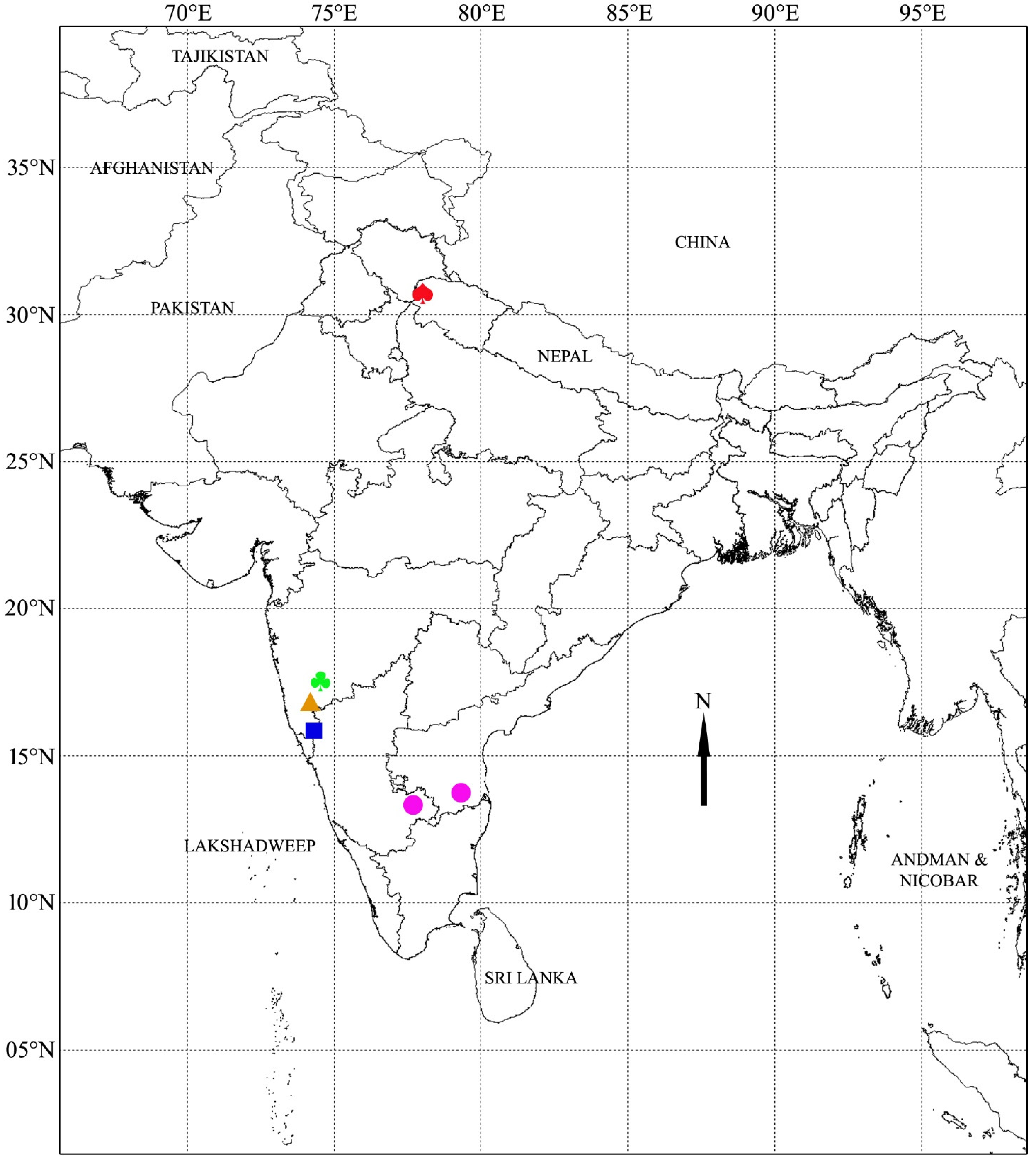
| Morphotype | Collection Locality | GPS Coordinates | Voucher ID | GenBank Accession Numbers | ||
|---|---|---|---|---|---|---|
| trnS-G | ndhF-trnL(UAG) | ITS | ||||
| Nandi Hills | ||||||
| B. cristata | Nandi Hills, Chikkaballapur district, Karnataka | N 13°21.400′ E 77°40.570′ | SSP 004 | ON667972 | ON693951 | ON706922 |
| B. cristata | Tirupati, Chittor district, Andhra Pradesh | N 13°41.157′ E 79°20.114′ | SSP 051 | ON667973 | ON693952 | ON706923 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 069 | ON667974 | ON693953 | ON706924 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 131 | ON667975 | ON693954 | ON706925 |
| Uttarakhand | ||||||
| B. cristata | Chakrata, Dehradun district, Uttarakhand | N 30°41.283′ E 77°52.215′ | SSP 023 | ON667976 | ON693955 | ON706926 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 055 | ON667977 | ON693956 | ON706927 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 065 | ON667978 | ON693957 | ON706928 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 134 | ON667979 | ON693958 | ON706929 |
| Cultivated form 1 | ||||||
| B. cristata | Kankumbi, Belgaum district, Karnataka | N 15°42.120′ E 74°13.130′ | SSP 024 | ON667980 | ON693959 | - |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 058 | ON667981 | ON693960 | - |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 079 | ON667982 | ON693961 | - |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 132 | ON667983 | ON693962 | - |
| Cultivated form 2 | ||||||
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 042 | ON667984 | ON693963 | ON706930 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 062 | ON667985 | ON693964 | ON706931 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 071 | ON667986 | ON693965 | ON706932 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 135 | ON667987 | ON693966 | ON706933 |
| Cultivated form 3 | ||||||
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 040 | ON667988 | ON693967 | ON706934 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 060 | ON667989 | ON693968 | ON706935 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 070 | ON667990 | ON693969 | ON706936 |
| B. cristata | Lead Botanical Garden, Shivaji University, Kolhapur, Maharashtra, India | N 16°40.310′ E 74°15.190′ | SSP 133 | ON667991 | ON693970 | ON706937 |
| Attributes | Barleria cristata (Cultivated form 1) | Barleria cristata (Cultivated form 2) | Barleria cristata (Cultivated form 3) | Barleria cristata (Nandi Hills) | Barleria cristata (Uttarakhand) |
|---|---|---|---|---|---|
| Leaves | Elliptic, 4.5–13.7 × 1.4–4.9 cm; venation eucamptodromous, one main vein from base, 5–8 pairs of principal laterals | Elliptic, 4–14.3 × 2–5.7 cm; venation eucamptodromous, one main vein from base, 5–8 pairs of principal laterals | Elliptic, 7.4–20.3 × 2.3–7.2 cm; venation eucamptodromous, one main vein from base, 5–7 pairs of principal laterals | Elliptic-lanceolate, 3.6–12.2 × 1.1–5.2 cm; venation eucamptodromous, one main vein from base, 6–10 pairs of principal laterals | Elliptic, 2–8.1 × 1–2.9 cm; venation eucamptodromous, one main vein from base, 4–5 pairs of principal laterals |
| Inflorescence | Cymes combining dichasial and monochasial elements with 1–6 flowers maturing sequentially | Cymes combining dichasial and monochasial elements with 1–7 flowers maturing sequentially | Cymes combining dichasial and monochasial elements with 1–8 flowers maturing sequentially | Unilateral cyme with 1–2 flowers maturing sequentially | Cymes combining dichasial and monochasial elements with 1–5 flowers maturing sequentially |
| Bracts | Linear, 0.8–1 × 0.1–0.15 cm, surface ciliate throughout, eglandular | Linear-lanceolate, 1.4–1.8 × 0.2–0.3 cm, surface ciliate throughout with stalked cupular glands near margins | Linear, 0.4–0.7 × 0.05–0.1 cm, surface with very few scattered stalked glandular trichomes | Linear-lanceolate, 1.4–1.8 × 0.2–0.3 cm, surface ciliate throughout with stalked cupular glandular trichomes near margins | Linear, 0.4–0.7 × 0.05–0.1 cm, surface ciliate throughout with very few scattered stalked glandular trichomes |
| Calyx | Outer lobes lance-ovate, 1.7–2.2 × 0.7–0.8 cm; apex narrowly acute, spinose; margins dentate; teeth 0.1–0.2 cm long, 7–12 | Outer lobes lance-ovate, 1.3–2.4 × 0.5–0.9 cm; apex narrowly acute, spinose; margins dentate; teeth 0.1–0.3 cm long, 8–15 | Outer lobes lance-ovate, 1.6–2.4 × 0.6–0.8 cm; apex narrowly acute, spinose; margins dentate; teeth 0.1–0.3 cm long, 8–15 | Outer lobes elliptic, 1.8–2.4 × 0.8–0.9 cm; apex acute, spinose; margins dentate; teeth 0.1–0.2 cm long, 16–22 | Outer lobes lance-ovate, 1.1–2.1 × 0.4–0.6 cm, apex narrowly acute, spinose; margins dentate; teeth 0.05–0.1 cm long, 9–11 |
| Inner lobes linear, 0.6–0.8 × 0.15–0.4 cm | Inner lobes linear, 0.6–0.8 × 0.15–0.2 cm | Inner lobes linear, 0.7–0.8 × 0.15–0.3 cm | Inner lobes linear, 0.6–0.9 × 0.1–0.2 cm | Inner lobes linear, 0.7–0.8 × 0.1 cm | |
| Corolla | Deep Rose Pink; upper lobes elliptic to broadly elliptic, apex acute or rounded; lower lobes broadly elliptic or suborbicular, apex retuse | White; upper lobes elliptic to broadly elliptic, apex acute or rounded; lower lobes broadly elliptic or suborbicular, apex rounded or acute | Lavender-violet; upper lobes lobes elliptic to broadly elliptic, apex acute or rounded; lower lobes very broadly elliptic or suborbicular, apex rounded or acute | Rose Pink; upper lobes obovate or oblanceovate, apex acute; lower lobes suborbicular or widely obovate, apex retuse | Chinese violet; upper lobes oblanceovate or elliptic, apex rounded or slightly retuse; lower lobes broadly elliptic or suborbicular, apex rounded |
| Stamens | 3–3.2 cm long; rose pink | 1.4–2.8 cm long; anthers lavender grey | 2.1–2.5 cm long; anthers lavender grey | 1.6–2.0 cm long; anthers wood browny | 1.5–2.6 cm long; anthers lavender grey |
| Gynoecium | Style 3.9–4.2 cm long, glabrous except near the base (few eglandular hairs) | Style 3.4–4.7 cm long, glabrous throughout | Style 3.2–4.6 cm long, glabrous throughout | Style 3.4–4.7 cm long, hirsute at base | Style 2.7–3.8 cm long, glabrous throughout |
| Morphotypes | P (µm) ± SD | E (µm) ± SD | P/E | Lumina Diameter (µm) ± SD | Colpus Length (µm) ± SD | Colpus Width (µm) ± SD | Pore Diameter (µm) ± SD |
|---|---|---|---|---|---|---|---|
| B. cristata (Cultivated form 1) | 79.79 ± 2.80 d | 83.51 ± 3.91 d | 0.96 a | 11.91 ± 2.32 b | 40.92 ± 3.46 b | 9.86 ± 1.24 d | 9.78 ± 1.13 c |
| B. cristata (Cultivated form 2) | 99.98 ± 4.16 a | 104.13 ± 4.02 a | 0.96 a | 17.39 ± 3.57 a | 38.23 ± 4.90 c | 11.50 ± 1.69 c | 8.51 ± 1.40 d |
| B. cristata (Cultivated form 3) | 100.73 ± 4.38 a | 104.13 ± 4.25 a | 0.97 a | 16.32 ± 4.25 a | 35.28 ± 4.61 d | 9.34 ± 1.60 d | 9.12 ± 1.14 bc |
| B. cristata (Nandi Hills) | 87.98 ± 4.49 c | 91.06 ± 4.69 c | 0.97 a | 13.15 ± 2.95 b | 47.28 ± 3.11 a | 16.50 ± 1.78 a | 11.93 ± 1.63 a |
| B. cristata (Uttarakhand) | 93.39 ± 4.16 b | 97.75 ± 4.19 b | 0.96 a | 15.42 ± 4.31 a | 39.63 ± 3.69 bc | 12.47 ± 1.29 b | 9.47 ± 1.21 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamboli, A.S.; Patil, S.S.; Kadam, S.K.; Choo, Y.-S.; Lekhak, M.M.; Pak, J.H. Morphology, Palynology and Molecular Phylogeny of Barleria cristata L. (Acanthaceae) Morphotypes from India. Diversity 2022, 14, 677. https://doi.org/10.3390/d14080677
Tamboli AS, Patil SS, Kadam SK, Choo Y-S, Lekhak MM, Pak JH. Morphology, Palynology and Molecular Phylogeny of Barleria cristata L. (Acanthaceae) Morphotypes from India. Diversity. 2022; 14(8):677. https://doi.org/10.3390/d14080677
Chicago/Turabian StyleTamboli, Asif S., Suraj S. Patil, Suhas K. Kadam, Yeon-Sik Choo, Manoj M. Lekhak, and Jae Hong Pak. 2022. "Morphology, Palynology and Molecular Phylogeny of Barleria cristata L. (Acanthaceae) Morphotypes from India" Diversity 14, no. 8: 677. https://doi.org/10.3390/d14080677