
Impacts of Diffuse Land-Use on Plant Diversity Patterns in the Miombo Woodlands of Western Zambia

1
Institute of Plant Science and Microbiology, Universität Hamburg, Ohnhorststr. 18, 22609 Hamburg, Germany
2
Compassionate Carbon Zambia Project, 3 Nangwenya Road, Rhodes Park, Lusaka 10101, Zambia
3
Institute for Globally Distributed Open Research and Education (IGDORE), Burgunderweg 9d, 22453 Hamburg, Germany
4
School of Medicine, Cavendish University, 47 Senanga Road, Handsworth Park, Lusaka P.O. Box 33145, Zambia
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(6), 739; https://doi.org/10.3390/d15060739
Submission received: 26 March 2023
/
Revised: 29 May 2023
/
Accepted: 29 May 2023
/
Published: 3 June 2023
(This article belongs to the Topic Diversity and Conservation of Flora in Africa)
Abstract
:Land use is known to influence the diversity of vascular plants in the Miombo woodlands. However, little is known about the interaction between soil and land use in herbaceous and woody species. We compared the diversity of vascular plants at the plot level (20 m × 50 m) and site level for three sites in the Miombo woodlands of western Zambia subject to different levels of intensity classes of diffuse land use (e.g., livestock herbivory and selective timber harvesting). For each of the sites, twenty plots were randomly selected for assessment of species composition of vascular plant species, indicators of land-use intensity, and soil chemistry per plot. We hypothesized that the site with the lowest human impact would have the highest richness and diversity of woody and herbaceous species. At the site level, we found that richness and diversity of woody species were unaffected by land-use intensity, whereas herbaceous species richness was higher for the protected site (28 species on average per 1000 m2) than the two other sites (23 and 21 species on average per 1000 m2). At the plot level, herbaceous species richness was positively associated with woodcutting and soil pH. We interpret the positive effect of woodcutting on herbaceous species richness as the effect of lower competition by the woody component for resources such as water, nutrients, and light. With regard to the absence of any effect of land-use intensity on the richness of woody species, we conclude that in our study areas selective timber harvesting may be at a sustainable level and might even have a positive effect on the diversity of the herbaceous layer.
1. Introduction
The Miombo woodlands form a widespread dry woodland belt covering large parts of southern and eastern Africa, encompassing Angola, Botswana, the Democratic Republic of Congo, Malawi, Mozambique, Tanzania, Zambia, and Zimbabwe [1]. While the flora of the Miombo woodlands is characterised by high species richness, the diversity of canopy trees is low [2]. The natural resources of the Miombo woodlands strongly contribute to the wealth of the social and economic systems in their distribution area [3]. This ecosystem and its resources, however, are prone to habitat transformation and biodiversity loss [4]. Anthropogenic activities such as fuel combustion, commercial selective harvesting of valuable timber, grazing, crop production, and complete deforestation threaten the woodland system [5,6,7]. With a deforestation rate of 1.5% per year, Zambia is even classified as one of the countries with the highest deforestation rates in the world [8]. Complete habitat conversion from forest to cropland inevitably decreases both herbaceous and woody species richness [9,10].
In contrast to deforestation, with its complete transformation of land cover, diffuse disturbance is characterised by relatively small patches of change distributed over a large area [11]. In woodlands, scattered, small-scale land use through grazing, selective timber harvesting, or small-scale cropping has a diffuse but continuous negative impact on the vegetation and woody and herbaceous forest biodiversity, as has been shown by studies of (sub)tropical forest ecosystems in southern Africa [12], Brazil [13], the mountains of Mexico [14], and the tropical rainforests of Madagascar [15]. Yet, the extent of the impact appears to vary from low to high [16,17]. Revermann et al. [12] showed how plant species richness and evenness respond to the diverging land-use patterns of spatially diffuse versus intense land use in the dry woodlands along the Kunene in Angola and Namibia. They showed that the spatially diffuse land use on unfenced communal land in Namibia has measurable negative effects on the richness of mainly woody plant species. The authors attributed this pattern to selective timber logging. In Zambia, Chidumayo [18] expressed a growing concern countrywide about the negative effects of diffuse disturbance due to the selective harvesting of trees for charcoal production and other uses. Irrespective of the potential impact of diffuse land use on botanical diversity, the effects of spatially-diffuse land use on the vegetation of the Miombo woodland have received little scientific attention in the available literature. Most of the researchers who have investigated the impact of land use on the Miombo woodlands have mainly focused on the effects on the woody component of the vegetation [3,19,20]. However, as has been shown by Revermann et al. [12], the response of forbs and grass species to diffuse disturbance of woodland systems may differ from that of woody species. The herbaceous layer of the Miombo woodlands shows great spatial variation in species composition, and several herbaceous genera contribute to the species richness and local endemism in the system [2]. Irrespective of the reported response of the herbaceous vegetation to diffuse land use in other vegetation types [12] and the strong contribution of herbaceous flora to the overall species richness and endemism, we are not aware of any study on the effect of land use on both the woody and herbaceous vegetation of the Miombo woodlands.
Therefore, in this study we focus on the effect of spatially diffuse land use on the composition and diversity of herbaceous and woody plant species in the western Zambian Miombo woodlands. To this end, we selected three sites in the Miombo woodlands of western Zambia, each representing a different land use type (i.e., national park, national forest, and community forest) related to a different level of spatially diffuse land-use intensity.
National Park (Kafue): no land-use activities are permissible, and only prescribed fires are used as a management intervention tool.
National Forest (Dongwe): a medium level of disturbance; only minimal land-use activities are allowed, and harvesting of timber is only allowed upon issuance of a permit.
Community Forest (Luampa): high level of disturbance with several land-use activities permitted, including the harvest of both timber and non-timber products as well as occasional agricultural activities.
We compared two species diversity measures for woody and herbaceous species at the three sites and analysed the spatially heterogeneous effects of indicators of diffuse land-use intensity and soil chemical variables on the species diversity measures at plot scale (1000 m2, n = 60).
Our hypothesis was that the Kafue site in the National Park would have a higher richness and diversity of woody and herbaceous species at both the plot and site scale than the two sites with higher land-use intensity. We additionally expected that, as observed by Revermann et al. [12], the diffuse disturbances would have a greater negative effect on species richness and biodiversity of trees than on the herbaceous layer.
2. Material and Methods
2.1. Study Area
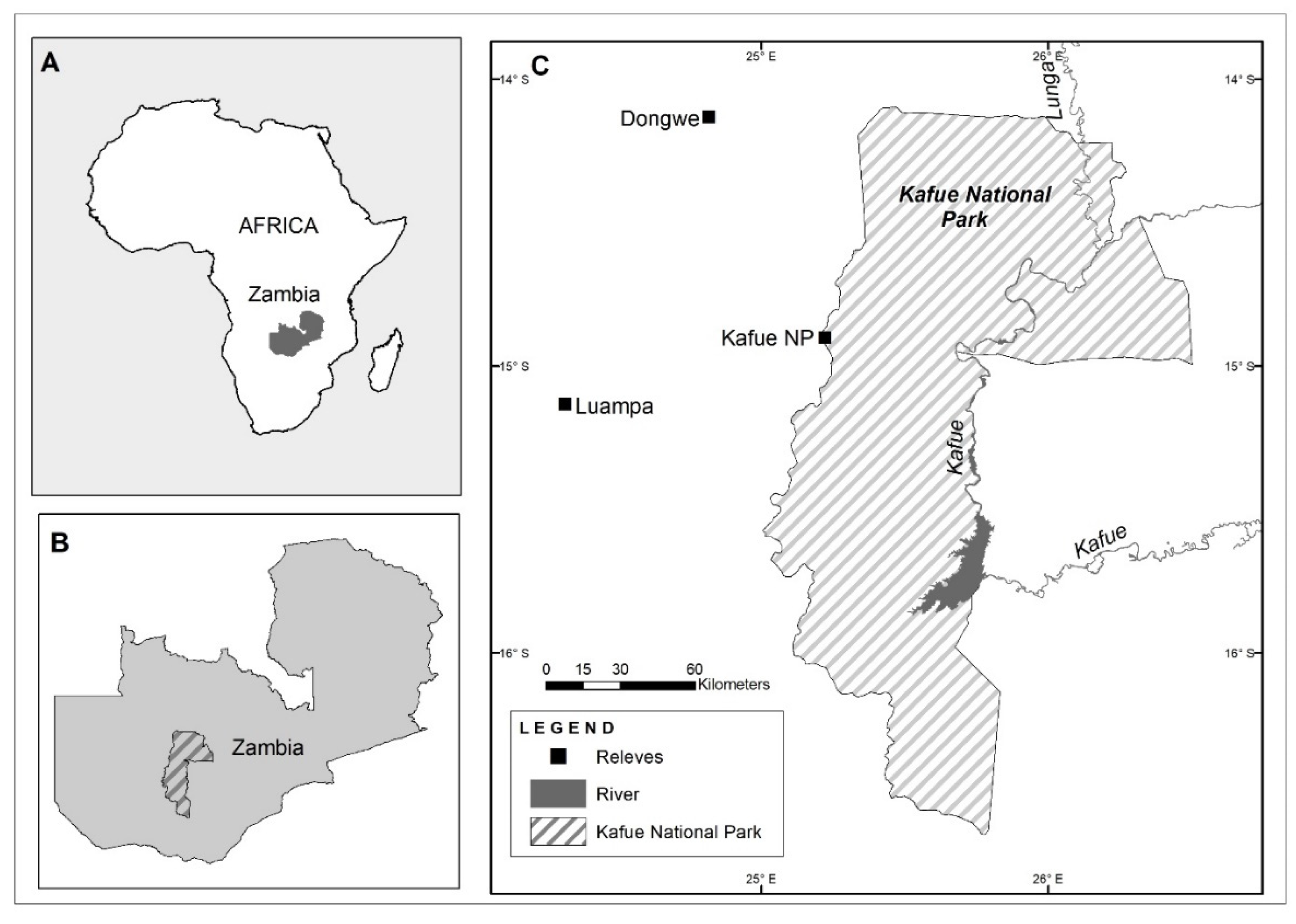
The study was conducted at three sites in the Miombo woodlands of western Zambia (Figure 1), extending over latitudes S 14°–16° and longitudes E 24°–26° at an elevation ranging from 1068 m to 1210 m a.s.l. (Table 1). The Miombo woodlands are the most extensive tropical seasonal woodland considered part of the African Savannah, covering about 2.4 million km2 in Africa [21]. Dominant plant families of Fabaceae tree species of the genera Brachystegia Benth., Isoberlina Craib and Stapf ex. Holland, and Julbernardia Pellegr., which are clustered in the subfamily Caesalpinioideae and the tribe Ahmerstieae [22,23,24], characterise the Miombo woodlands [24]. The area has a tropical sub-humid climate with alternating dry and wet seasons. Rainfall occurs for 5–7 months in summer [25]. The mean annual temperature of the study area is 20.8 °C, and the mean annual rainfall ranges from 875 to 990 mm (Table 1). The soils consist of Kalahari sands from the Tertiary to the recent period which, according to Japan Association for International Collaboration of Agriculture and Forestry [26], cover western and northwestern Zambia. The main soil type is Arenosols, a formation of the parent Basement and Katanga rocks, with the accumulation of Karoo deposits [27].
Floristically, the three sites fall within the Sudano-Zambezian Phytoregion [28], whereas the vegetation is classified as Miombo woodlands sensu [29], which forms part of the tropical seasonal woodlands [30]. Miombo woodlands are characterized by co-dominance of trees, shrubs, and herbaceous plants, with respective proportions determined by environmental, ecological, and human parameters and adapted to fire [31]. The dominant tree species are those referred to by Phiri [24] as dominant taxa of the Miombo woodlands.
2.2. Study Design and Data Collection
We selected three study sites in Kafue National Park (called Kafue for the rest of the text), Dongwe, and Luampa, approximately 100 km apart (Figure 1), based on literature consultations [29,32] and site visits to the study area of western Zambia in 2014. Classification according to land-use type and intensity was guided by the State of the Environment in Zambia [33]. Each site was characterised by different land-use intensities, from low to medium to high (Table 1). For each study site, we classified the vegetation into open or closed woodland using the most recent Google Earth image [34] at the time of data assessment. Based on habitat stratification, we randomly selected twenty vegetation plots for each of the sites. The plots were 1000 m2 (20 m × 50 m) in size and were laid out in an east–west extension.

{kind=link}
{kind=link}
Table 1.
Descriptive attributes of the three study sites.
| Site Name | Mean Annual Rainfall [mm/yr] | Mean Annual Temperature [°C] | Land-Use Type | Land-Use Intensity | GPS Coordinates (North West Corner) | Elevation Min–Max [masl] |
|---|---|---|---|---|---|---|
| Luampa | 875.5 | 22.22 | Forest Reserve | High | S15.13782, E24.48778 | 1152–1158 |
| Dongwe | 990.6 | 22.34 | Community Forest | Medium | S14.09577, E24.01520 | 1066–1145 |
| Kafue NP | 897.4 | 21.98 | National Park | Low | S14.89830, E25.43676 | 1091–1210 |
For each of the plots, we recorded the presence and identity of all angiosperms as herbaceous or woody species (as defined by Petruzzello [37], with the former being plants that do not have a true woody stem and may be either perennial or annual), estimated cover per species in percent, and counts of individuals per species (see Appendix A, Table A1 for all recorded species identified). Grass species had to be excluded from the analyses because at the time of sampling the majority of grasses did not have inflorescence, which compromised identification of the species. To assess the presence of exotic species and field weeds in the study area, we reviewed the literature sources in Zambia [38,39,40,41]. Indicators for land-use intensity, such as signs of recent woodcutting, grazing, and browsing, were recorded semi-quantitatively. The categories of woodcutting were 1–2 stumps per 1000 m2-plot = 1, 3–5 stumps = 2, 6–8 stumps = 3, and >8 stumps = 4. Signs of browsing from game animals (domestic livestock were not observed in any of the study sites) were graded from 1 (no browsing) to 4 (high abundance of signs of browsing) depending on the frequency of signs of browsing observed. Grazing was not observed on any of the plots. The time from the last fire event on the plot was determined based on consultation with local field assistants as well as our own observations of the age of visible signs. We did not determine the cause of the fire. However, in areas where human activity is frequent, fires caused by human activity are more common than natural causes [42]. The determined time from the last fire was translated to the ordinal scale for the recent occurrence of fire: long ago = 2 (10 or more years since last fire), recent = 1 (5 years), and very recent = 0 (1 or 2 years).
One composite soil sample comprising five subsamples per plot was collected from the topsoil layer (0–10 cm). Soil samples were analysed at the Mt. Makulu Research Centre Soil Laboratory, Chilanga, Zambia for the variables of pH, nitrogen (N), phosphorus (P), organic carbon (Org C), calcium (Ca), magnesium (Mg), sodium (Na), potassium (K), zinc (Zn), manganese (Mn), iron (Fe), and cation exchange capacity (CEC). For the soil analytical methods, see Appendix A, Table A2.
Within the plots, vegetation data were collected during the rainy seasons (March–May) of 2014 and 2015. The data obtained from the Zambia Meteorology Department (ZMD) indicated that the annual rainfall in these years was at 50–70% of the mean annual rainfall, which ranges between 875 and 990 mm for the study area (Table 1). Mean annual rainfall per site was available for the period from 1950 to 2010 from the Global Land Data Assimilation System site [35].
Plant species identification in the field was carried out using field guides [43,44,45,46] and later confirmed at the herbarium of the University of Zambia. Plant nomenclature followed Phiri [24] for the majority of the identifications; in cases of ambiguity, the Flora of Zambia [39], the JSTOR Global Plants database [47], and the Plant List [48] were used for verification. Voucher specimens were lodged at the Herbarium Hamburgense of the Universität Hamburg (HBG) and the herbarium of the University of Zambia (UZL).
2.3. Data Analysis
Species inventories were analysed for richness (S) and Shannon index per plot [49]. The sites differed in the degree of exposure to human activities (Table 1). We tested the semi-quantitative land-use variables (browsing, grazing, woodcutting, and time from last fire) for differences among the three land-use intensities by applying a Kruskal–Wallis test and then running a pairwise comparison of the land-use variables.
We tested the normal distribution of the diversity data. For the non-normally distributed Shannon diversity values, which are already on a log scale, we used the exponential of the Shannon, which is a common way to transform Shannon entropy into Shannon diversity values [50]. The diversity values were then expressed as the effective number of species. We applied one-way ANOVA to test for differences between the species richness values and the exponential of the Shannon under different land-use intensities followed by a Tukey HSD post hoc test.
We were interested in how well land-use intensity and soil chemistry can explain variation in richness and Shannon diversity of woody and herbaceous plants at the plot scale across the three study sites. We screened all soil variables visually for skewed distributions, which are common for these kinds of data. Of the twelve measured soil variables, we had to log-transform eight (P, Ca, Mg, K, Na, Zn, Mn, Fe). We tested the environmental variables for pairwise correlation to exclude multicollinearity, and only maintained the relevant variables of fire, woodcutting, browsing, soil pH, and above-mentioned eight soil variables. We employed generalised linear models (GLM) using the Poisson distribution, as the data were discrete and had a lower boundary (zero) for richness. A Gaussian distribution was used for the exponential of the Shannon diversity values. For the four response variables in the GLM (i.e., the richness and the diversity of woody and herbaceous species, respectively), we created a full model including soil and land-use variables as predictors. We used a heuristic approach that compared all 2100 possible models (not using interactions) and identified a final best model. All models were fitted using the R package glmulti [51]. Because we were using GLMs, we used a ‘pseudo’-R2 measure, which behaves like R2 and measures the improvement of a fitted model compared to a null model calculated as the ratio [52]. Here, we used Cragg and Uhler’s R2 available from the R package pscl [53].
3. Results
3.1. Indicators for Land-Use Intensities at the Three Sites
We compared the indicator values for diffuse land-use intensity among the three sites. Table 2 shows that the two sites which were subject to diffuse anthropogenic disturbance, Dongwe National Forest and Luampa Community Forest, showed more frequent signs of woodcutting and shorter time from the last fire along with a lower frequency of signs of browsing compared to the Kafue site.
3.2. Diversity Patterns at the Different Land-Use Intensities
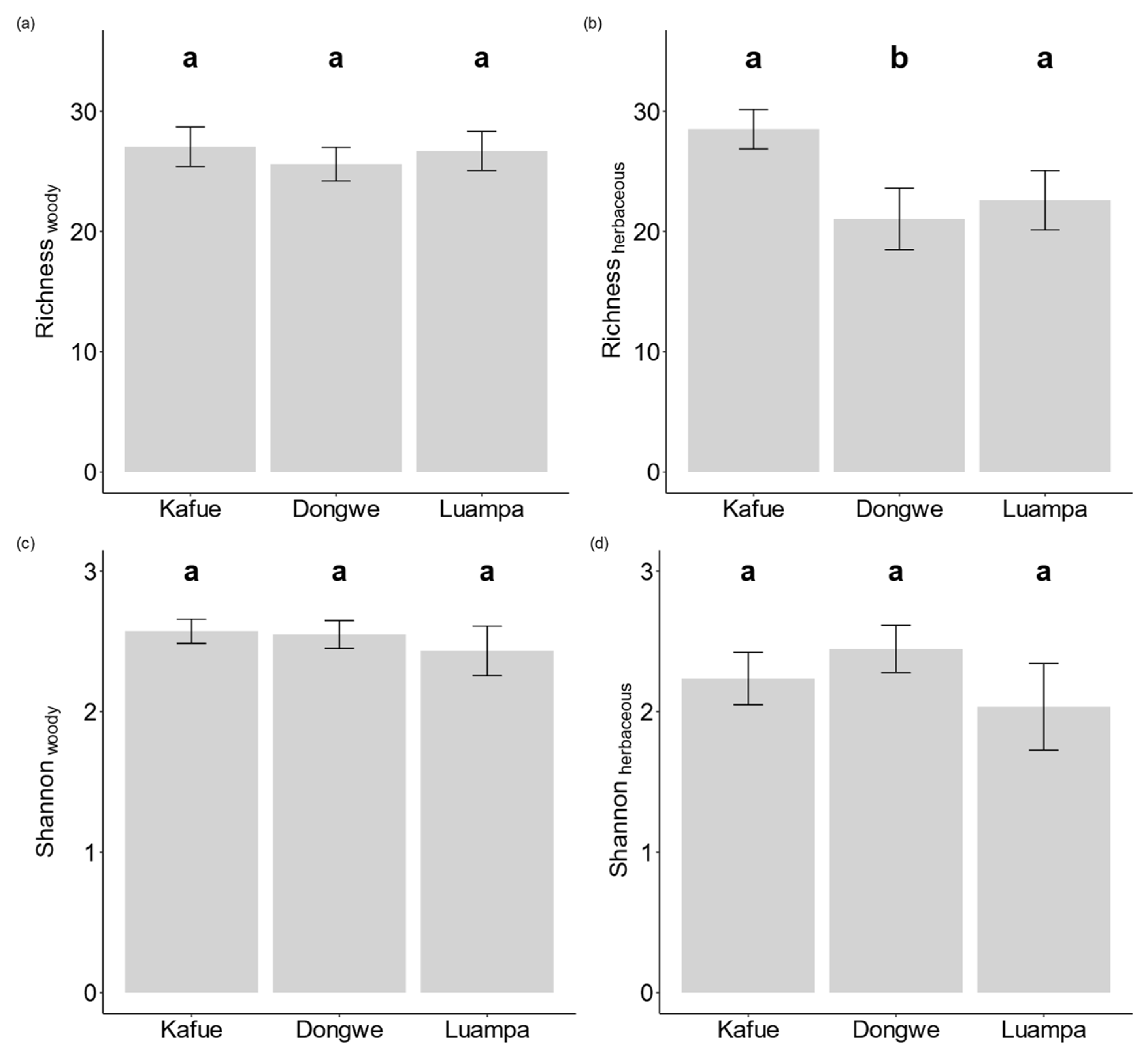
Due to our pre-classification of the sites into three different land-use types (Table 1) and the higher frequency of recorded signs of anthropogenic disturbances at the two sites under human land use (Luampa and Dongwe) compared with the National Park site (Kafue, Table 2), we expected strong differences between the diversity indicators in the sites. For all three sites, a cumulative total of 624 vascular plant species from 51 plant families was recorded. Of these species, 239 were woody and 385 were herbaceous. The mean species richness of the twenty plots (1000 m2) per site for woody species was 27 (Luampa), 25 (Dongwe), and 27 (Kafue), while for herbaceous species it was 23 (Luampa), 21 (Dongwe), and 28 (Kafue) (Figure 2). In contrast to our expectations, species richness and Shannon diversity at the plot level only differed in terms of herbaceous species (Figure 2). Kafue had on average eight more herbaceous species per 1000 m2 plot than the other two sites. The mean richness of woody species per plot was about 26 species, and the mean diversity was at about 13 effective species at all three sites. Very little research has been published on exotic and invasive plant species in woodland habitats [38,39,40,41]. Of the herbaceous species observed in the semi-disturbed sites of our study (Luampa and Dongwe), the following herbaceous species have been classified as weeds in the literature: Crassocephalum rubens, Striga asiatica, Vernonia petersii, and Crotalaria spp. Dichrostachys cineria, a species reported by Blaser-Hart et al. [40] to cause bush encroachment in Zambia, was observed in the woody vegetation of all the sites, though with low density.
3.3. Environmental Drivers of Diversity at Plot Level
Although the recorded disturbance variables of woodcutting, browsing, and fire events differed between Kafue and the other two sites, their distribution varied within each of the sites as well (Table 2). We were therefore interested in the effects of the plot-based land-use intensities on the diversity measures per plot. We related the observed signs of disturbance per plot to the species richness and Shannon diversity of woody and herbaceous plant species for the respective plots. To discriminate between the disturbance effects and those of other abiotic habitat variables, we included plot-based soil variables in the analyses as well. For each of the four response variables (i.e., richness and diversity of woody and herbaceous species), we did not find any significant variation in richness or diversity of the woody component in response to any of the disturbance or soil variables (Table 3). We did, however, find a positive response of the species richness in the herbaceous layer to an increase in soil pH and to woodcutting at the plot level. The Shannon diversity of herbaceous species solely responded to organic carbon (positively) and iron (negatively). Overall, the models were rather weak, with low pseudo-R2 values except for the richness of the herbaceous layer.
4. Discussion
4.1. Effect of Land Use on Plant Species Diversity
Our study showed that the two sites under medium and high land-use-intensity, namely, Dongwe Forest Reserve and Luampa Community Forest, indeed had higher frequency of woodcutting and a shorter time from the last fire than Kafue (Table 2). Selective woodcutting in the Miombo woodlands in Zambia targets valuable timber species. The most harvested species according to the Forest Department are Baikiaea plurijuga and Pterocarpus angolensis [54], which are used for both domestic and commercial purposes. Selective harvesting of valuable timber species could lead to overharvesting and even local extinction, as has been shown by De Cauwer et al. [55] for Pterocarpus angolensis in Namibia. Therefore, we expected woodcutting for selective timber harvesting to have a negative effect on woody species richness and diversity. Even though intensity of woodcutting intensity differed between Kafue and the other two sites, woody species richness and diversity were not significantly different among the three sites. One possible reason for this might be that the observed woodcutting intensity at Dongwe Forest Reserve and Luampa Community Forest, with 0–3 tree stumps per 1000 m2, while significantly higher than in Kafue (Table 2), was too low to have a measurable negative effect on the tested diversity measures. In previous studies, very moderate land-use intensity did not show any negative effect on species richness and abundance of woody species in the Miombo woodlands of Tanzania [56] and in the West African savannah of Burkina Faso [57], and even showed an increase in the tree species diversity of the Miombo woodlands in Angola [12]. It appears that in the present study the impact of land use on woody species diversity was below the threshold, in contrast to that observed in other studies [56,57]. This observation shows that the offtake of woody species through woodcutting and destruction by fire can be considered sustainable if the impact remains on a small spatial scale.
In contrast to the woody species richness and diversity, we found the richness of herbaceous species to be lower at Luampa and Dongwe than at the Kafue National Park site, suggesting a negative impact of land use on the herbaceous layer. Nacoulma et al. [57] compared the herbaceous species richness of sites under land use and in protected areas for savannah woodlands in Burkina Faso. They found that sites under higher land-use intensity to have lower herbaceous species richness, which they explained by the effect of higher grazing intensity in the unprotected area. Unsustainable grazing pressure from livestock reduces the number of palatable herbaceous species in woodlands [58]. The impact of grazing livestock (i.e., livestock that predominantly consume the herbaceous layer of the vegetation) on biodiversity has been addressed by previous studies on savannah ecosystems [59,60,61], showing that grazing may, depending on its intensity, have either a positive or negative effect on species richness. In our study, however, we did not find any signs of grazing in either Kafue, where one might expect game to graze, or at either of the other two sites, where grazing domestic cattle could be expected. The observed absence of grazing signs at the Miombo woodland site in Kafue supports reports that the grazers of the national park prefer the open grasslands (called dambos) [62]. Similarly, at Luampa and Dongwe, where grazing is also absent, the livestock farmers prefer to graze their cattle in the open grasslands (dambos), where the quality of forage is better.
Although we did not find any signs of grazing (consumption of the vegetation of the herbaceous layer), we did find signs of browsing (consumption of the leaves on twigs and branches of shrubs and trees), which was significantly less at the two sites under land use (Luampa and Dongwe) than at Kafue. At Luampa and Dongwe, browsing livestock such as goats have been restricted to areas close to land users’ homesteads, and are kept away from the distant woodlands, according to personal communication with local farmers. At Kafue, we found signs of browsing, even though no presence or signs of large mammals were observed. The browsing signs could be attributed to the presence of insects known to browse on woody species [62]. However, increased density of browsing insect species in the woodlands of Kafue National Park is unlikely to have a negative effect on the woody component of the vegetation. This means that the low density of woody species at the Kafue site cannot be explained by grazing. Reduced woody cover facilitates the abundance of herbaceous species by reducing the competition with woody components for resources such as light, water, and nutrients [30].
Field weeds and invasive species such as Crassocephalum rubens, Striga asiatica, Vernonia petersii, and Crotalaria spp. were recorded only occasionally or even rarely in the plots of the semi-disturbed study sites. Dichrostachys cineria was only rarely observed at all three sites. Owing to the very limited information available in the literature on exotic and invasive plant species in the woodlands of Zambia, very few species in our study were identified as exotic or invasive, and these had very low densities. Therefore, our data suggest that the influence of synanthropic plants on the diversity patterns is likely to be low.
4.2. Drivers of Diversity at Plot Level
The range of land-use variables per site (Table 2) revealed within-site variability of land-use effects within the three study sites. Therefore, we tested for the effect of land-use intensity on species richness and diversity per plot. Because soil characteristics show interplay with biotic drivers and drive vascular plant diversity in the Miombo woodlands [25], we tested the effect of both land-use indicators and soil variables on diversity and richness at plot level irrespective of the land-use type they were exposed to. Land-use (woodcutting) and soil variables (soil pH) showed effects on the diversity patterns of herbaceous species, whereas woody species diversity and richness were not affected. The absence of land-use effects on woody species diversity and richness is in contrast to other studies showing that selective harvesting of valuable timber species can lead to local extinction [55]. The patchy nature of diffuse disturbance of woodlands referred to by Asefa et al. [63] is defined by the absence of settlements and crop cultivation as well as relatively low-level logging and grazing, and had positive effects on woody species richness in Ethiopia. In turn, we found the herbaceous species richness to increase with woodcutting. As discussed earlier, this positive effect might have been a result of better light conditions for the herbaceous species coupled with reduced competition for space, water, and nutrient resources [4,30]. Reduction of competition for resources in combination with the release of organic nutrients through decaying tree stumps have previously been shown to increase herbaceous biomass in the Miombo woodlands [64]. At the site level, Kafue had the highest herbaceous species richness as well as the highest density in terms of signs of browsing (Table 2). Browsing, as discussed earlier, may already have a positive effect on herbaceous species and species diversity. Our study showed a positive effect of diffuse disturbance caused by woodcutting (plot level) and browsing (site level in Kafue) on herbaceous species richness and diversity.
We further found a positive relationship between richness of herbaceous species and an increase in soil pH at the plot level. Soils in humid subtropical regions are typically acidic [65]. In our study area, the dominant soil type (Arenosols) is leached under high rainfall conditions, resulting in low pH [66], with soil pH ranging from 3.7 to 5.4 (Appendix A, Table A3). Acidity in soils slows down the rate of decomposition of soil organic material, and thereby reduces the availability of nutrients for plant uptake [67]. Thus, highly acidic soils may provide a low nutrient supply, which limits the range of plant species that can cope with these conditions [68]. Therefore, very low soil pH has previously been found to be negatively associated with species diversity of herbaceous species in the Miombo woodlands [2,69,70] and the tropical montane forests of Cameroon [71].
We further found SOC at the plot level to be negatively associated with the Shannon diversity of herbaceous species. Generally, SOC content increases with precipitation and with optimal levels in humid and cold climates and decreases with soil pH; beyond that, SOC storage links to biophysical factors and management practices [72]. A study in Ghana by Quaye et al. [73] revealed unsuitably low SOC in strongly acidic soils in the western African Savannah woodlands. In the Miombo Woodlands of our study area, which are Savannah woodlands, low soil pH might have negatively affected SOC as well. The SOC appeared to be negatively related to the biodiversity of herbaceous species, which could be because herbaceous species have a lower root network than woody species [30].
At the plot level, our study showed that both land use (woodcutting) and soil acidity were drivers of the diversity of herbaceous species, whereas woody species were unaffected. The absence of variance in the richness and diversity of woody species, however, does not exclude the fact that there are differences in other vegetation characteristics, such as species composition. Variances in small-scale species composition in response to fire and herbivory have previously been shown in the Miombo woodlands of Zimbabwe [74]. We expect similar effects in our study area, which we will analyse in a subsequent study.
5. Conclusions
This study revealed that diffuse land use has no influence on woody species richness and diversity in the Miombo woodlands in our study area. The absence of such effects on woody species richness and diversity was consistent across all three of our sites. However, the study showed influence of both diffuse land use and soil acidity on herbaceous species richness at the plot level. Among the land-use parameters, woodcutting (at plot level) and browsing (at site level) showed positive effects on herbaceous species richness; both of these result in opening up of the tree canopy, providing water, nutrients, and sunlight for herbaceous species. Other land-use effects, such as fire and grazing, which have been shown in other studies to influence patterns of richness and diversity of vascular plant species in the region, were not found in our study. We assume that the intensities of these disturbances at all three sites and plots were too low to show any effects.
Author Contributions
P.S. (Hamburg University)—Data sampling, data analysis, and drafting of the manuscript; J.O. (Hamburg University)—Statistical support; N.J. (Hamburg University)—Study design and conceptual guidance; P.P. (Cavendish University)—Identification of specimens and conceptual guidance; U.S. (Hamburg University)—Study design, conceptual guidance, and support in drafting the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was part of the BMBF-funded SASSCAL initiative (Task no. 159) promotion number 01LG1201M.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data in this publication is deposited in BIOTABase and can be accessed on request from [email protected].
Acknowledgments
This study formed a part of the research portfolio of the BMBF-funded SASSCAL initiative (Task no. 159) promotion number 01LG1201M. We are very grateful to the field assistants who participated in the collection of the data, namely, Alex Liseli, Andrew Munshukulumbwe, and Charles Sitali. Gerhard Muche and Rasmus Revermann provided guidance for the use of the database software BIOTABase and R statistics package. We are grateful for the support on climate data provided by Verena Baumberg. Zambia Wildlife Authority granted us permission to undertake this study in its protected areas.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
List of woody and herbaceous plant species with their life form, collection number, and location, arranged according to family: Abundance was assigned according to the frequency of observations of each woody plant species: rare (1 or 2 recordings), occasional (3–5 recordings), frequent (6–10 recordings), and common (>11 recordings). The locations where these species occurred in the study sites are abbreviated as D = Dongwe, K = Kafue National Park, and L = Luampa. The different uses were coded as TI = timber production, PO = posts, pole, and roundwood, WO = fuelwood and charcoal, PU = pulp and paper production, FD = fodder, FO = food, NW = other non-wood products (gums, medicines, dyes, tanning, etc.), AE = aesthetic and ethical values, and TX = toxic to livestock.
Table A1.
List of woody and herbaceous plant species with their life form, collection number, and location, arranged according to family: Abundance was assigned according to the frequency of observations of each woody plant species: rare (1 or 2 recordings), occasional (3–5 recordings), frequent (6–10 recordings), and common (>11 recordings). The locations where these species occurred in the study sites are abbreviated as D = Dongwe, K = Kafue National Park, and L = Luampa. The different uses were coded as TI = timber production, PO = posts, pole, and roundwood, WO = fuelwood and charcoal, PU = pulp and paper production, FD = fodder, FO = food, NW = other non-wood products (gums, medicines, dyes, tanning, etc.), AE = aesthetic and ethical values, and TX = toxic to livestock.
| Species Name | Life Form | Abundance | Uses | Voucher Number | Location |
|---|---|---|---|---|---|
| Acanthaceae Duosperma quadrangulare (Klotzsch) Brummitt Hypoestes forskaolii (Vahl) R.Br. | |||||
| Herbaceous | rare | FO | 138076 | K | |
| Herbaceous | TX | 132148 | K | ||
| Amaryllidaceae Crinum macowanii Baker | rare | ||||
| Herbaceous | rare | NW | 132141 | L, K | |
| Anacardiaceae | |||||
| Searsia quartiniana (A. Rich.) A.J. Mill. | Woody | Frequent | FO | 131071, 142196 | L, D |
| Sclerocarya birrea (A.Rich.) Hochst. | Woody | rare | FO | 132128 | K |
| Annonaceae | |||||
| Friesodielsia obovata (Benth.) Verdc. | Woody | occasional | FO, FD | 142610 | K |
| Uvariastrum hexaloboides (R.E.Fr.) | Woody | occasional | FO | 132107 | L, K |
| Xylopia odoratissima Welw. ex Oiv. | Woody | common | NW | 131181 | L, D |
| Apocynaceae | |||||
| Diplorhynchus condylocarpon (Müll. Arg.) Pichon | Woody | common | NW | 140121 | D, L, K |
| Landolphia parvifolia K. Schum. | Woody | frequent | FO | 142505 | D, L |
| Strophanthus welwitschii (Baill.) K. Schum. | Woody | occasional | 142649 | D | |
| Asparagaceae | |||||
| Asparagus racemosus Willd. | Herbaceous | occasional | 142788 | K | |
| Asphodelaceae | |||||
| Bulbine abyssinica A.Rich. | Herbaceous | rare | NW | 131355, 132214 | D, K |
| Asteraceae | |||||
| Conyza gouanii (L.) Willd. | Herbaceous | occasional | NW | 132207 | K |
| Crassocephalum rubens (Jacq.) S. Moore | Herbaceous | rare | NW | 131301 | D |
| Dicoma anomala Sond. | Herbaceous | occasional | 138099 | K | |
| Elephantopus scaber L. | Herbaceous | frequent | 131119 | D, L, K | |
| Erythrocephalum zambesianum Oliv. & Hiern | Herbaceous | frequent | NW | 142503 | L, K |
| Felicia welwitschii (Hiern) Grau | Herbaceous | rare | 142698 | D | |
| Macledium poggei (O.Hoffm.) S.Ortiz | Herbaceous | rare | 142660 | K | |
| Pleiotaxis eximia O. Hoffm. Vernonia glabra ssp. laxa (Seetz) Vatke | Herbaceous | occasional | NW | 142665 | L, K L, K |
| Vernonia melleri Oliv. & Hiern | Herbaceous | occasional | 131401 | L | |
| Vernonia petersii Oliv. & Hiern ex Oliv. | Herbaceous | frequent | FD | 142678 | D, K |
| Vernonia poskeana Vatke & Hildebr. | Herbaceous | rare | 131157 | L | |
| Capparaceae | |||||
| Capparis tomentosa Lam. | Herbaceous | frequent | NW, FO, AE | 138093 | K |
| Cleome hirta (Klotzsch) Oliv. | Herbaceous | frequent | 138076 | K | |
| Maerua triphylla ssp. pubescens A. Rich. (Klotzsch) DeWolf | Herbaceous | frequent | 131358 | K | |
| Chrysobalanaceae | |||||
| Parinari capensis Harv. | Woody | frequent | NW, FO | L | |
| Parinari curatellifolia Planch. ex Benth. | Woody | frequent | NW, FO, WO | 131213 | L, K |
| Combretaceae | |||||
| Combretum collinum Fresen. | Woody | occasional | NW, TI | 131113 | D, L, K |
| Combretum elaeagnoides Klotzsch | Woody | occasional | 131403 | D, L | |
| Combretum molle R.Br. ex G.Don | Woody | frequent | NW, TI | 142578 | D, L, K |
| Combretum psidioides Welw. | Woody | occasional | NW | 131402 | D |
| Combretum zeyheri Sond. | Woody | common | NW, WO | 131183 | L, K |
| Pteleopsis anisoptera (Welw. ex M.A.Lawson) Engl. & Diels | Woody | occasional | NW, TI, WO | 1320102 | D |
| Terminalia brachystemma Welw. ex Hiern | Woody | occasional | NW, FO, WO | 131165 | L, K |
| Commelinaceae | |||||
| Cyanotis longifolia Benth. | Herbaceous | frequent | 131016 | D, L, K | |
| Dipterocarpaceae | |||||
| Marquesia macroura Gilg | Woody | occasional | 140121 | K | |
| Gilg | Woody | occasional | 132222 | K | |
| Monotes glaber Sprague | Woody | occasional | NW, WO | 142639, 141189 | L |
| Ebenaceae | |||||
| Diospyros batocana Hiern | Woody | common | NW, FO, PU | 140159 | D, L, K |
| Diospyros mespiliformis Hochst. ex A.DC. | Woody | frequent | NW, FO | K | |
| Diospyros virgata (Gürke) Brenan | Woody | common | NW | 131054, 131191, 140143 | D, L, K |
| Ericaceae | |||||
| Cleistanthus polystachyus Hook. f. ex Planch. | Herbaceous | rare | 132122 | L | |
| Erythroxylaceae | |||||
| Erythroxylum emarginatum Thonn. | Woody | occasional | 142649 | K | |
| Euphorbiaceae | |||||
| Acalypha ornata Hochst. ex A. Rich. | Herbaceous | occasional | 132101 | L | |
| Flueggea virosa (Roxb. ex Willd.) Voigt | Woody | occasional | NW, FO | 142566 | L, K |
| Hymenocardia acida Tul. | Woody | common | NW | 142526 | D, L, K |
| Maprounea africana Müll.Arg. | Woody | frequent | NW | L | |
| Oldfieldia dactylophylla (Welw. ex Oliv.) Léonard | Woody | frequent | 142693 | D | |
| Pseudolachnostylis maprouneifolia Pax | Woody | common | NW, FO | 140131, 131047 | D, L, K |
| Sclerocroton oblongifolius (Müll. Arg.) Kruijt & Roebers | Herbaceous | rare | NW | 131056, 142172, 135679 | L, K |
| Uapaca kirkiana Müll. Arg. | Woody | occasional | NW, FO | 142817 | D, K |
| Uapaca nitida ssp. nitida | Woody | occasional | NW, FO | L, K | |
| Fabaceae | |||||
| Afzelia quanzensis Welw. | Woody | frequent | NW, FO TI, WO | 131163 | L, K |
| Albizia antunesiana Harms | Woody | frequent | NW, WO | 142766 | D, L, K |
| Albizia versicolor Welw. ex Oliv. | Woody | common | NW, WO | 132156 | D, K |
| Anisophyllea boehmii Engl. | Woody | occasional | NW, FO | 132216 | K |
| Baphia massaiensis var. obovata Taub. | Woody | common | NW, FO, FD | 131007 | D, L, K |
| Bauhinia petersiana Bolle | Woody | frequent | NW, FO, FD | 142520 | D, L, K |
| Bobgunnia madagascariensis (Desv.) J.H.Kirkbr. & Wiersema | Woody | common | NW, FO, TI | 131191 | L, K |
| Brachystegia boehmii Taub. | Woody | common | TI, WO, PO, NW | D, L, K | |
| Brachystegia spiciformis Benth. | Woody | common | TI, WO, PO, NW | 131146 | D, L, K |
| Burkea africana Hook. | Woody | common | WO, PO, NW | D, L, K | |
| Cassia abbreviata Oliv. | Woody | occasional | NW | 142622 | K |
| Chamaecrista mimosoides (L.) Greene | Herbaceous | occasional | NW | 131025, 142559 | D, K |
| Copaifera baumiana Harms | Woody | common | NW | 131085 | L, K |
| Crotalaria alexandri Baker f. | Herbaceous | occasional | 142576 | L, D | |
| Crotalaria anisophylla (Hiern) Welw. ex Baker f. | Herbaceous | occasional | 142774 | L | |
| Crotalaria caudata Welw. ex Baker | Herbaceous | occasional | 131176 | D, L | |
| Crotalaria cephalotes Steud. ex A.Rich. | Herbaceous | occasional | NW | 132180 | K |
| Crotalaria laburnifolia L. | Herbaceous | common | 142570 | D | |
| Crotalaria microcarpa Hochst. ex Benth. | Herbaceous | rare | 131099 | D, L | |
| Cryptosepalum exfoliatum ssp. pseudotaxus De Wild. | Woody | common | TI, NW, PO, | 142504 | D |
| Dichrostachys cinerea (L.) Wight & Arn. | Woody | common | NW, WO | D, L, K | |
| Erythrophleum africanum | Woody | common | WO, PO, NW | 131124 | D, L, K |
| Guibourtia coleosperma (Benth.) J.Leonard | Woody | common | TI, NW, | 140151 | D, L |
| Indigofera demissa Taub. | Herbaceous | occasional | 131174 | D, L | |
| Indigofera flavicans Baker | Herbaceous | common | 131130 | L, K | |
| Isoberlinia angolensis (Benth.) Hoyle & Brenan | Woody | occasional | TI, NW, AE | K | |
| Julbernardia paniculata (Benth.) Troupin | Woody | common | TI, WO, PO, NW, FD | 140155 | D, L, K |
| Keetia venosa (Oliv.) Bridson | Woody | occasional | 142561 | K | |
| Lannea edulis (Sond.) Engl. | Woody | frequent | NW | 131178 | L, K |
| Mucuna poggei Taub. | Woody | occasional | NW | 142564 | K |
| Pericopsis angolensis (Baker) Meeuwen | Woody | occasional | TI, WO, NW, FD | 138072 | L, K |
| Piliostigma thonningii (Schumach.) Milne-Redh. | Woody | occasional | K | ||
| Pterocarpus angolensis DC. | Woody | common | TI, WO, NW | D, L, K | |
| Rhynchosia caribaea (Jacq.) DC. | Woody | occasional | NW | 131195 | L, K |
| Flacourtiaceae | |||||
| Flacourtia indica (Burm.f.) Merr. | Woody | frequent | NW, FO | 131198 | K |
| Hypericaceae | |||||
| Psorospermum baumii Engl. | Woody | occasional | NW, FO | 131162 | D |
| Lamiaceae | |||||
| Ocimum africanum Lour. | Herbaceous | rare | K | ||
| Tinnea vestita Baker | Herbaceous | occasional | NW, FO | 131021, 142739 | L, K |
| Vitex doniana Sweet | Woody | occasional | TI, FO, NW, FD | 140154 | L |
| Vitex madiensis Oliv. subsp. milanjiensis (Britten) F. White | Woody | rare | 142638 | L | |
| Lauraceae | |||||
| Cassytha pondoensis ssp. Pondoensis Engl. | Woody | occasional | NW | 131134 | D, L |
| Malvaceae | |||||
| Abutilon angulatum (Guill. & Perr.) Mast. | Woody | occasional | NW | 142504 | D |
| Pavonia senegalensis (Cav.) Leistner | Woody | rare | 131268 | D, L | |
| Meliaceae | |||||
| Bersama abyssinica Fresen. | Woody | rare | NW, AE | 142874 | K |
| Trichilia emetica Vahl | Woody | rare | NW, AE | K | |
| Myrtaceae | |||||
| Syzygium guineense (Willd.) DC. | Woody | occasional | NW, FO | 142200 | L |
| Ochna pulchra Hook. | Woody | common | NW | 131059 | D, L, K |
| Olacaceae | |||||
| Olax obtusifolia De Wild. | Woody | occasional | 142598 | L | |
| Ximenia americana L. | Woody | frequent | NW, FO | 142523 | L, K |
| Ximenia caffra Sond. | Woody | frequent | NW, FO | 131047 | K |
| Oleaceae | |||||
| Olea capensis L. | Woody | occasional | 138065 | K | |
| Schrebera trichoclada Welw. | Woody | rare | 144524 | L, K | |
| Orobanchaceae | |||||
| Striga asiatica (L.) Kuntze | Herbaceous | occasional | NW | 138077 | L, D |
| Oxalidaceae | |||||
| Biophytum abyssinicum Steud. Ex A. Rich. | Herbaceous | occasional | NW | D | |
| Biophytum umbraculum Welw. | Herbaceous | occasional | 131098 | D, L | |
| Passifloraceae | |||||
| Paropsia brazzeana Baill. | Woody | frequent | NW | 131080 | D, L, K |
| Polygalaceae | |||||
| Securidaca longepedunculata Fresen. | Woody | occasional | NW | 131204 | L |
| Proteaceae | |||||
| Protea angolensis Welw. | Woody | rare | NW | 142795 | K |
| Protea gaguedi J.F. Gmel. | Woody | frequent | NW | K | |
| Ranunculaceae | |||||
| Clematis chrysocarpa Welw. ex Oliv. | Herbaceous | occasional | 131351, 142755, 142609 | L, K | |
| Rhamnaceae | |||||
| Ziziphus mucronata Willd. | Woody | occasional | FO, NW | K | |
| Rubiaceae | |||||
| Agathisanthemum bojeri Klotzsch | Woody | occasional | NW | 142781 | L, K |
| Fadogia cienkowskii Schweinf. | Woody | occasional | 142191 | L | |
| Gardenia ternifolia Schumach. & Thonn. | Woody | occasional | NW, FO | 132132 | D |
| Pavetta schumanniana F. Hoffm ex K. Schum | Woody | frequent | 142749 | D, K | |
| Rothmannia engleriana (K.Schum.) Keay | Woody | frequent | NW | 131196 | D, L |
| Spermacoce pusilla Wall. | Woody | occasional | 131100 | D, L | |
| Tricalysia longituba De Wild. | Woody | occasional | 131255 | L, K | |
| Vangueriopsis lanciflora (Hiern) Robyns | Woody | frequent | NW, WO | 131180 | D, L |
| Sapindaceae | |||||
| Zanha africana (Radlk.) Exell | Woody | occasional | WO | 142751 | K |
| Sapotaceae | |||||
| Englerophytum magalismontanum (Sond.) T.D.Penn. | Herbaceous | occasional | NW | 131079 | D, L |
| Solanaceae | |||||
| Solanum mauritianum Scop. | Herbaceous | occasional | K | ||
| Strychnaceae | |||||
| Strychnos cocculoides Baker | Woody | frequent | NW, FO, AE, PO | 131083 | D, L, K |
| Strychnos pungens Soler. | Woody | frequent | NW, FO | 140149 | D, L, K |
| Thelypteridaceae | |||||
| Christella chaseana (Schelpe) Holttum | Herbaceous | occasional | D | ||
| Tiliaceae | |||||
| Grewia flavescens Juss. | Herbaceous | occasional | NW, FO | 131013 | D, L, K |
| Triumfetta annua L. | Herbaceous | occasional | 131109 | L, K | |
| Verbenaceae | |||||
| Endostemon obtusifolius (E. Mey. ex Benth.) N.E.Br. | Herbaceous | occasional | D, L | ||
| Lantana angolensis Moldenke | Herbaceous | occasional | NW, FO, FD | 142644 | L, K |
| Vitaceae | |||||
| Cyphostemma junceum Wild & R.B. Drumm. | Herbaceous | frequent | NW, FO | 132169 | D |
| Cyphostemma princeae Wild & R.B. Drumm | Herbaceous | frequent | NW, FO | 131382 | D, L, K |
| Zingiberaceae | occasional | ||||
| Aframomum alboviolaceum (Ridl.) K.Schum. | Herbaceous | NW, FO | D |
Table A2.
Methods of soil sample analysis.
| Analysis Variable | Method and Reference | Unit |
|---|---|---|
| pH | using the CaCl2 mixture in H2O [75] | |
| Nitrogen (N) | Kjeldahl method [76] | percentage per total weight |
| Phosphorus (P) | Bray I extractant [77] | parts per million |
| Organic Carbon (Org C) | Walkley Black technique [78] | percentage per total weight |
| Calcium (Ca) | Ammonia acetate extraction [79] | parts per million |
| Magnesium (Mg) | Ammonia acetate extraction [79] | parts per million |
| Sodium (Na) | Ammonia acetate extraction [79] | parts per million |
| Potassium (K) | Ammonia acetate extraction [79] | parts per million |
| Zinc (Zn) | DPTA method [80] | parts per million |
| Manganese (Mn) | DPTA method [80] | parts per million |
| Iron (Fe) | DPTA method [80] | parts per million |
| Cation Electronic Exchange (CEC) | Conductivity method [81] | Milli-equivalents |
Table A3.
Soil variables from the observatory samples analysis.
| Variables | Mean and Covariance | Luampa | Dongwe | Kafue National Park |
|---|---|---|---|---|
| pH | 4.4 | 3.9 | 4.9 | |
| CV | 0.04 | 0.07 | 0.08 | |
| N | 0.016 | 0.011 | 0.018 | |
| CV | 0.5 | 0.64 | 0.66 | |
| P | 5.0381 | 3.01 | 7.73 | |
| CV | 0.42 | 0.39 | 0.83 | |
| Org C | 0.25 | 0.21 | 0.26 | |
| CV | 0.43 | 0.37 | 0.5 | |
| Ca | 40 | 32 | 120 | |
| CV | 0.71 | 1.61 | 0.49 | |
| Mg | 10.23 | 11 | 24.31 | |
| CV | 0.58 | 0.14 | 0.57 | |
| Na | 2.23 | 3.75 | 6.53 | |
| CV | 1.49 | 0.64 | 0.65 | |
| K | 10.91 | 10.35 | 42.05 | |
| CV | 1.28 | 1.14 | 0.48 | |
| Zn | 0.27 | 0.05 | 0.15 | |
| CV | 2.19 | 1.43 | 1.4 | |
| Mn | 14.42 | 12.1 | 82.05 | |
| CV | 1.15 | 2.6 | 0.58 | |
| Fe | 11.23 | 11.95 | 28.47 | |
| CV | 0.49 | 0.84 | 0.51 | |
| CEC | 3.45 | 2.4 | 3.83 | |
| CV | 0.35 | 0.58 | 0.32 |
Soil variables with for mean values and CV for covariance.
References
- Campbell, B.M.; Angelsen, A.; Cunningham, A.B.; Katerere, Y.; Sitoe, A.; Wunder, S. Miombo Woodlands—Opportunities and Barriers to Sustainable Forest Management; Centre for International Forest Research: Bogor, Indonesia, 2008. [Google Scholar]
- Frost, P. The ecology of miombo woodlands. In The Miombo in Transition: Woodlands and Welfare in Africa; Campbell, B., Ed.; Centre for International Forestry Research: Bogor, Indonesia, 1996; pp. 11–57. [Google Scholar]
- Kalaba, F.K.; Quinn, C.H.; Dougill, A.J.; Vinya, R. Floristic composition, species diversity and carbon storage in charcoal and agriculture fallows and management implications in Miombo woodlands of Zambia. For. Ecol. Manag. 2013, 304, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Chidumayo, E.N.; Gumbo, D.J. The Dry Forests and Woodlands of Africa: Managing for Products and Services; Chidumayo, E.N., Gumbo, D.J., Eds.; Centre for International Forest Research: London, UK, 2010. [Google Scholar]
- Chapin, F.S., 3rd; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Wessels, K.J.; Reyers, B.; Van Jaarsveld, A.S. Incorporating land cover information into regional biodiversity assessments in South Africa. Anim. Conserv. 2000, 3, 67–79. [Google Scholar] [CrossRef]
- Walters, D.J.J.; Kotze, D.C.; O’connor, T.G. Impact of land use on vegetation composition, diversity, and selected soil properties of wetlands in the southern Drakensberg mountains, South Africa. Wetl. Ecol. Manag. 2006, 14, 329–348. [Google Scholar] [CrossRef]
- Vinya, R.; Syampungani, S.; Kasumu, C.E.; Monde, C.; Kasubika, R. Preliminary Study on the Drivers of Deforestation and Potential for REDD+ in Zambia. A Consultancy Report Prepared for Forestry Department and FAO under the National UN-REDD+ Programme; Ministry of Lands & Natural Resources: Lusaka, Zambia, 2011. [Google Scholar]
- Hawthorne, W.D.; Marshall, C.A.M.; Abu-Juam, M.; Agyeman, V.K. The Impact of Logging Damage on Tropical Rain-Forests, Their Recovery and Regeneration: An Annotated Bibliography; Oxford Forestry Institute: Oxford, UK, 2011; 123p. [Google Scholar]
- Newbold, T.; Tittensor, D.P.; Harfoot, M.B.J.; Scharlemann, J.P.W.; Purves, D.W. Nonlinear changes in modelled terrestrial ecosystems subjected to perturbations. Sci. Rep. 2020, 10, a14051. [Google Scholar] [CrossRef]
- White, P.S.; Jentsch, A. The Search for Generality in Studies of Disturbance and Ecosystem Dynamics. Botany 2001, 62, 399–450. [Google Scholar] [CrossRef]
- Revermann, R.; Wallenfang, J.; Oldeland, J.; Finckh, M. Species richness and evenness respond to diverging land-use patterns—A cross-border study of dry tropical woodlands in southern Africa. Afr. J. Ecol. 2016, 55, 152–161. [Google Scholar] [CrossRef]
- Barlow, J.; Lennox, G.D.; Ferreira, J.; Berenguer, E.; Lees, A.C.; Mac Nally, R.; Thomson, J.R.; Ferraz, S.F.D.B.; Louzada, J.; Oliveira, V.H.F.; et al. Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature 2016, 535, 144–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Marcial, N.; González-Espinosa, M.; Williams-Linera, G. Anthropogenic disturbance and tree diversity in Montane Rain Forests in Chiapas, Mexico. For. Ecol. Manag. 2001, 154, 311–326. [Google Scholar] [CrossRef]
- Vallan, D.; Andreone, F.; Raherisoa, V.H.; Dolch, R. Does selective wood exploitation affect amphibian diversity? The case of An’Ala, a tropical rainforest in eastern Madagascar. Oryx 2004, 38, 410–417. [Google Scholar] [CrossRef] [Green Version]
- Savadogo, P.; Tiveau, D.; Sawadogo, L.; Tigabu, M. Herbaceous species responses to long-term effects of prescribed fire, grazing and selective tree cutting in the savanna-woodlands of West Africa. Perspect. Plant Ecol. Evol. Syst. 2008, 10, 179–195. [Google Scholar] [CrossRef]
- Kalema, V.N.; Witkowski, E.T. Land-use impacts on woody plant density and diversity in an African savanna charcoal production region. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2012, 8, 231–247. [Google Scholar] [CrossRef] [Green Version]
- Chidumayo, E.N. Species structure in Zambian miombo woodland. J. Trop. Ecol. 1987, 3, 109–118. [Google Scholar] [CrossRef]
- Chidumayo, E.N. Changes in miombo woodland structure under different land tenure and use systems in central Zambia. J. Biogeogr. 2002, 29, 1619–1626. [Google Scholar] [CrossRef]
- Zisadza-Gandiwa, P.; Mabika, C.T.; Kupika, O.L.; Gandiwa, E.; Murungweni, C. Vegetation Structure and Composition across Different Land Uses in a Semiarid Savanna of Southern Zimbabwe. Int. J. Biodivers. 2013, 2013, 692564. [Google Scholar] [CrossRef]
- Dewees, P.A.; Campbell, B.M.; Katerere, Y.; Sitoe, A.; Cunningham, A.B.; Angelsen, A.; Wunder, S. Managing the Miombo woodlands of Southern Africa: Policies, incentives and options for the rural poor. J. Nat. Resour. Policy Res. 2010, 2, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.G. Ecological Results of Woodland and Burning Experiments in Northern Rhodisia. J. Ecol. 1959, 47, 129. [Google Scholar] [CrossRef]
- Werger, M.J.A.; Coetzee, B.J. The Sudano-Zambezian region. In Biogeography and Ecology of Southern Africa; Monographiae Biologicae; Springer: Dordrecht, The Netherlands, 1978. [Google Scholar] [CrossRef]
- Phiri, P. A Checklist of Zambian Vascular Plants; Southern African Botanical Diversity Network: Pretoria, South Africa, 2005. [Google Scholar]
- Desanker, P.V.; Frost, P.G.H.; Frost, C.O.; Justice, C.O.; Scholes, R.J. (Eds.) The Miombo Network: Framework for a Terrestrial Transect Study of Land-Use and Land-Cover Change in the Miombo Ecosystems of Central Africa; IGBP Report 41; The International Geosphere-Biosphere Programme (IGBP): Stockholm, Sweden, 1997; 109p. [Google Scholar]
- Japan Association for International Collaboration of Agriculture and Forestry. Agriculture and Forestry in Zambia: Present Situation and Issues for Future Development; Japan Association for International Collaboration of Agriculture and Forestry: Tokyo, Japan, 2008. [Google Scholar]
- Survey Department. Soil map of Zambia; Survey Department: Lusaka, Zambia, 1983. [Google Scholar]
- White, F. The savannah woodlands of the Zambesian and Sudanian domain: An ecological and phytogeographical comparison. Syst. Assn. Publ. 1965, 4, 71–103. [Google Scholar]
- White, F. The Vegetation of Africa: A Descriptive; United Nations Educational Scientific and Cultural Organisation: Paris, France, 1983. [Google Scholar]
- Hofiço, N.D.S.A.; Fleig, F.D. Diversity and Structure of Miombo Woodlands in Mozambique Using a Range of Sampling Sizes. J. Agric. Sci. Technol. B 2015, 5, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Furley, P.A.; Rees, R.M.; Ryan, C.M.; Saiz, G. Savanna burning and the assessment of long-term fire experiments with particular reference to Zimbabwe. Prog. Phys. Geogr. Earth Environ. 2008, 32, 611–634. [Google Scholar] [CrossRef] [Green Version]
- Chidumayo, E.N. Miombo Ecology and Management: An Introduction; IT Publications in association with the Stockholm Environment Institute: London, UK, 1997; 166p. [Google Scholar]
- Environmental Council of Zambia. State of Environment Report in Zambia 2000; Environmental Council of Zambia: Lusaka, Zambia, 2001. [Google Scholar]
- Google Earth 6.0–Version. Western Province, Zambia. 14040′51.22″ S, 24043′14.54″ E Eye Alt 225.46 mi. Digital Globe 2012. 2014. Available online: http://www.earth.google.com (accessed on 17 April 2014).
- GLDAS. Global Land Data Assimilation System. GLDAS NOAA R0.25 deg 3H 1948–2010. 2014. Available online: https://developers.google.com/earth-engine/datasets/catalog/NASA_GLDAS_V021_NOAH_G025_T3H (accessed on 12 May 2015).
- Ministry of Tourism Environment and Natural Resources. Current Protected Area Categories and the Institutions Responsible for Their Management in Zambia; Ministry of Tourism Environment and Natural Resources: Lusaka, Zambia, 2004. [Google Scholar]
- Petruzzello, M. Herbaceous Plant. Encyclopedia Britannica, 17 January 2023. 2023. Available online: https://www.britannica.com/plant/herbaceous-plant (accessed on 19 April 2023).
- Vernon, R. Field Guide to Important Arable Weeds of Zambia; Department of Agriculture: Chilanga, Zambia, 1983; 45p. [Google Scholar]
- Bingham, M.; Willemen, A.; Wursten, B.; Ballings, P.; Hyde, M. Flora of Zambia. Lists of Families. 2011. Available online: http://www.zambiaflora.com/speciesdata/listoffamilies (accessed on 16 March 2016).
- Blaser, W.J.; Shanungu, G.K.; Edwards, P.J.; Venterink, H.O. Woody encroachment reduces nutrient limitation and promotes soil carbon sequestration. Ecol. Evol. 2014, 4, 1423–1438. [Google Scholar] [CrossRef] [PubMed]
- Siachoono, S.M.; Syampungani, S.; Mundike, J. Invasive Alien Species in Zambia. Invasive Alien Species 2021, 1, 313–329. [Google Scholar] [CrossRef]
- Nieman, W.A.; Van Wilgen, B.W.; Leslie, A.J. A review of fire management practices in African savanna-protected areas. Koedoe 2021, 63, 13. [Google Scholar] [CrossRef]
- Storrs, A. Know Your Trees—Some of the Common Trees Found in Zambia; Forest Department: Kitwe, Zambia, 1979. [Google Scholar]
- Klassen, E.S.; Craven, P. Checklist of grasses in Namibia. Southern African Botanical Diversity Network Report No. 20; SABONET: Windhoek, Namibia, 2003. [Google Scholar]
- Le Roux, P.; Mueller, M. Le Roux and Müller’s Field Guide to the Trees and Shrubs of Namibia; Macmillan Education: Windhoek, Namibia, 2009. [Google Scholar]
- van Wyk, B.; van Wyk, P. Field Guide to Trees of Southern Africa: An African Perspective; Struik Publishers: Cape Town, South Africa, 2013. [Google Scholar]
- JSTOR. Global Plants Database. Available online: https://plants.jstor.org/ (accessed on 16 March 2016).
- The Plant List. 2013. Available online: http://www.theplantlist.org/ (accessed on 23 April 2016).
- Scheiner, S.M. Biological Diversity: Frontiers in Measurement and Assessment. Edited by Anne E. Magurran and Brian J. McGill. Oxford and New York: Oxford University Press. $135.00 (hardcover); $72.50 (paper). xvii + 345 p.; ill.; index. ISBN: 978-0-19-958066-8 (hc); 978-0-19-958067-5 (pb). 2011. Q. Rev. Biol. 2012, 87, 254. [Google Scholar] [CrossRef]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Calcagno, V.; Mazancourt, C. de glmulti: An R Package for Easy Automated Model Selection with (Generalized) Linear Models. J. Stat. Softw. 2010, 34, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Long, S.J. Regression Models for Categorical and Limited Dependent Variables; Sage Publications: Thousand Oaks, CA, USA, 1997. [Google Scholar]
- Jackman, S.; Tahk, A.; Zeileis, A.; Maimone, C.; Fearon, J.; Meers, Z. Package pscl: Classes and Methods for R Developed in the Political Science Computational Laboratory; CRAN: Stanford, CA, USA, 2017; pp. 1–97. [Google Scholar]
- Ng’andwe, P.; Mwitwa, J.; Muimba-Kankolongo, A. Forest Policy, Economics, and Markets in Zambia; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1–166. [Google Scholar]
- De Cauwer, V.; Muys, B.; Revermann, R.; Trabucco, A. Potential, realised, future distribution and environmental suitability for Pterocarpus angolensis DC in southern Africa. For. Ecol. Manag. 2014, 315, 211–226. [Google Scholar] [CrossRef]
- Jew, E.K.; Dougill, A.J.; Sallu, S.M.; O’connell, J.; Benton, T.G. Miombo woodland under threat: Consequences for tree diversity and carbon storage. For. Ecol. Manag. 2016, 361, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Nacoulma, B.M.I.; Schumann, K.; Traoré, S.; Bernhardt-Römermann, M.; Hahn, K.; Wittig, R.; Thiombiano, A. Impacts of land-use on West African savanna vegetation: A comparison between protected and communal area in Burkina Faso. Biodivers. Conserv. 2011, 20, 3341–3362. [Google Scholar] [CrossRef]
- Smet, M.; Ward, D. A comparison of the effects of different rangeland management systems on plant species composition, diversity and vegetation structure in a semi-arid savanna. Afr. J. Range Forage Sci. 2005, 22, 59–71. [Google Scholar] [CrossRef]
- Giliba, R.A.; Boon, E.K.; Kayombo, C.J.; Musamba, E.B.; Kashindye, A.M.; Shayo, P.F. Species Composition, Richness and Diversity in Miombo Woodland of Bereku Forest Reserve, Tanzania. J. Biodivers. 2011, 2, 1–7. [Google Scholar] [CrossRef]
- Hanke, W.; Boehner, J.; Dreber, N.; Juergens, N.; Schmiedel, U.; Wesuls, D.; Dengler, J. The impact of livestock grazing on plant diversity: An analysis across dryland ecosystems and scales in southern Africa. Ecol. Appl. 2014, 24, 1188–1203. [Google Scholar] [CrossRef] [Green Version]
- Odadi, W.O.; Fargione, J.; Rubenstein, D.I. Vegetation, Wildlife, and Livestock Responses to Planned Grazing Management in an African Pastoral Landscape. Land Degrad. Dev. 2017, 28, 2030–2038. [Google Scholar] [CrossRef]
- Zambia Wildlife Authority. Kafue National Park General Management Plan 2011–2020; Zambia Wildlife Authority: Lusaka, Zambia, 2010. [Google Scholar]
- Asefa, A.; Yosef, M.; Mengesha, G.; Mamo, Y.; Shimelis, A. Woody plant diversity along disturbance gradients in the Northern Afro-Montane Forests of the Bale Mountains, Ethiopia. Int. J. Dev. Res. 2015, 5, 3745–3754. [Google Scholar]
- Shirima, D.D.; Pfeifer, M.; Platts, P.; Totland, Ø.; Moe, S.R. Interactions between Canopy Structure and Herbaceous Biomass along Environmental Gradients in Moist Forest and Dry Miombo Woodland of Tanzania. PLoS ONE 2015, 10, e0142784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, A.D.; Ludeke, K.L. Plant Nutrients in Desert Environments; Springer: Berlin, Germany, 1993. [Google Scholar] [CrossRef]
- Silva, M.M.V.G.; Cabral-Pinto, M.M.S.; Dinis, P. Geochemistry of subtropical arenosols from Kuito region (Angola). Urbanization effects and environmental implications. J. Afr. Earth Sci. 2021, 183, 104307. [Google Scholar] [CrossRef]
- McCauley, A.; Jones, C.; Olson-Rutz, K. Soil pH and organic matter. Nutr. Manag. Modul. 2017, 8, 1–12. [Google Scholar]
- Kamelarczyk, K.B.F. Carbon Stock Assessment and Modelling in Zambia A UN-REDD Programme Study; National Forest Resource and Assessment Programme: Lusaka, Zambia, 2009. [Google Scholar]
- Shackleton, C.; Shackleton, S.E.; Buiten, E.; Bird, N. The importance of dry woodlands and forests in rural livelihoods and poverty alleviation in South Africa. For. Policy Econ. 2007, 9, 558–577. [Google Scholar] [CrossRef]
- Chidumayo, E. Forest degradation and recovery in a miombo woodland landscape in Zambia: 22 years of observations on permanent sample plots. For. Ecol. Manag. 2013, 291, 154–161. [Google Scholar] [CrossRef]
- Fonge, B.A.; Tchetcha, D.J.; Nkembi, L. Diversity, Distribution, and Abundance of Plants in Lewoh-Lebang in the Lebialem Highlands of Southwestern Cameroon. Int. J. Biodivers. 2013, 2013, 642579. [Google Scholar] [CrossRef] [Green Version]
- Marks, E.; Aflakpui, G.K.S.; Nkem, J.; Poch, R.M.; Khouma, M.; Kokou, K.; Sagoe, R.; Sebastià, M.-T. Conservation of soil organic carbon, biodiversity and the provision of other ecosystem services along climatic gradients in West Africa. Biogeosciences 2009, 6, 1825–1838. [Google Scholar] [CrossRef] [Green Version]
- Quaye, A.K.; Doe, E.K.; Attua, E.M.; Yiran, G.; Arthur, A.; Dogbatse, J.A.; Konlan, S.; Nkroma, Y.D.; Addo, D. Geospatial distribution of soil organic carbon and soil pH within the cocoa agroecological zones of Ghana. Geoderma 2021, 386, 114921. [Google Scholar] [CrossRef]
- Mapaure, I. Small-Scale Variations in Species Composition of Miombo Woodland in Sengwa, Zimbabwe: The Influence of Edaphic Factors, Fire and Elephant Herbivory. Syst. Geogr. Plants 2001, 71, 935. [Google Scholar] [CrossRef]
- Adams, F.; Evans, C.E. A Rapid Method for Measuring Lime Requirement of Red-Yellow Podzolic Soils. Soil Sci. Soc. Am. J. 1962, 26, 355–357. [Google Scholar] [CrossRef]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Anal. Bioanal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Chapman, H.D. Total Exchangeable Bases. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; John Wiley: Hoboken, NJ, USA, 1965; pp. 902–904. [Google Scholar] [CrossRef]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Chapman, H.D.; Wilcox, L.V. Soluble salts. In Methods of Soil Analysis; Norman, A.G., Ed.; Soil Science Society of America: Madison, WI, USA, 1965. [Google Scholar] [CrossRef]
Figure 1.
Locations of study sites (C) in western Zambia, showing the sites in relation to the continent (A) and the country (B).
Figure 1.
Locations of study sites (C) in western Zambia, showing the sites in relation to the continent (A) and the country (B).

Figure 2.
Bar plots of the diversity indices of the 1000 m2 plots for species richness and Shannon index as follows (a) Richness for the woody species; (b) Richness for the herbaceous species; (c) Shannon index for the woody species and; (d) Shannon index for the herbaceous species. Different groups (based on Tukey’s HSD test) are indicated by superscripts.
Figure 2.
Bar plots of the diversity indices of the 1000 m2 plots for species richness and Shannon index as follows (a) Richness for the woody species; (b) Richness for the herbaceous species; (c) Shannon index for the woody species and; (d) Shannon index for the herbaceous species. Different groups (based on Tukey’s HSD test) are indicated by superscripts.

Table 2.
Pairwise comparisons using the Wilcoxon Rank Sum Test. Median values for the levels of the land-use variables. Figures in brackets = the range of the observed intensity of the respective land-use variables; hyperscripts indicate differences between sites at a significance level of p < 0.05.
Table 2.
Pairwise comparisons using the Wilcoxon Rank Sum Test. Median values for the levels of the land-use variables. Figures in brackets = the range of the observed intensity of the respective land-use variables; hyperscripts indicate differences between sites at a significance level of p < 0.05.
| Land-Use | Luampa (High Land-Use Intensity) [Median Values and (in Brackets) Min–Max Values] | Dongwe (Medium Land-Use Intensity) [Median Values and (in Brackets) Min–Max Values] | Kafue (No Land-Use) [Median Values and (in Brackets) Min–Max Values] |
|---|---|---|---|
| Woodcutting | 1 a (0–3) | 1 a (0–3) | 0 b (0–1) |
| Browsing | 0 a (0–1) | 0 a (0–1) | 1 b (0–2) |
| Time since last fire | 0 a (0–2) | 1 a (0–2) | 2 b (1–2) |
Table 3.
Generalised linear models for species richness (S) and the exponential of the Shannon diversity (H’) of woody and herbaceous species after multi-model inference. The values for the logarithm of iron (logFe) and woodcutting are chi-square statistics, indicating an overall significant effect of the parameters. pR2 is Cragg and Uhler’s pseudo r-squared [51] measure based on the differences between best model and null model. SOC = soil organic carbon.
Table 3.
Generalised linear models for species richness (S) and the exponential of the Shannon diversity (H’) of woody and herbaceous species after multi-model inference. The values for the logarithm of iron (logFe) and woodcutting are chi-square statistics, indicating an overall significant effect of the parameters. pR2 is Cragg and Uhler’s pseudo r-squared [51] measure based on the differences between best model and null model. SOC = soil organic carbon.
| Diversity Model | Distribution and Link | n | Intercept | logMg | SOC | pH | logFe ‡ | Wood Cutting ‡ | pR2 |
|---|---|---|---|---|---|---|---|---|---|
| Swoody | Poisson (log) | 60 | 3.275 *** | 0.00 | |||||
| Sherbs | Poisson (log) | 60 | 2.593 *** | 0.151 ** | 9.432 * | 0.36 | |||
| H’woody | Gaussian (id) | 60 | 11.880 *** | 0.935 | −5.970 | 0.07 | |||
| H’herbs | Gaussian (id) | 60 | 2.773 *** | 2.506 | −12.952 * | −2.141 * | 0.18 |
* p < 0.05; ** p < 0.01; *** p < 0.001 ‡ Chi-square statistic.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sichone, P.; Oldeland, J.; Phiri, P.; Jürgens, N.; Schmiedel, U. Impacts of Diffuse Land-Use on Plant Diversity Patterns in the Miombo Woodlands of Western Zambia. Diversity 2023, 15, 739. https://doi.org/10.3390/d15060739
AMA Style
Sichone P, Oldeland J, Phiri P, Jürgens N, Schmiedel U. Impacts of Diffuse Land-Use on Plant Diversity Patterns in the Miombo Woodlands of Western Zambia. Diversity. 2023; 15(6):739. https://doi.org/10.3390/d15060739
Chicago/Turabian StyleSichone, Priscilla, Jens Oldeland, Patrick Phiri, Norbert Jürgens, and Ute Schmiedel. 2023. "Impacts of Diffuse Land-Use on Plant Diversity Patterns in the Miombo Woodlands of Western Zambia" Diversity 15, no. 6: 739. https://doi.org/10.3390/d15060739
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.