Multiple Factors Influence Seasonal and Interannual Litterfall Production in a Tropical Dry Forest in Mexico

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Selection of Sampling Sites
2.3. Estimation of Litterfall Production
2.4. Climatic Variables
2.5. Measurement and Estimation of Vegetation Variables
2.6. Data Analysis
3. Results
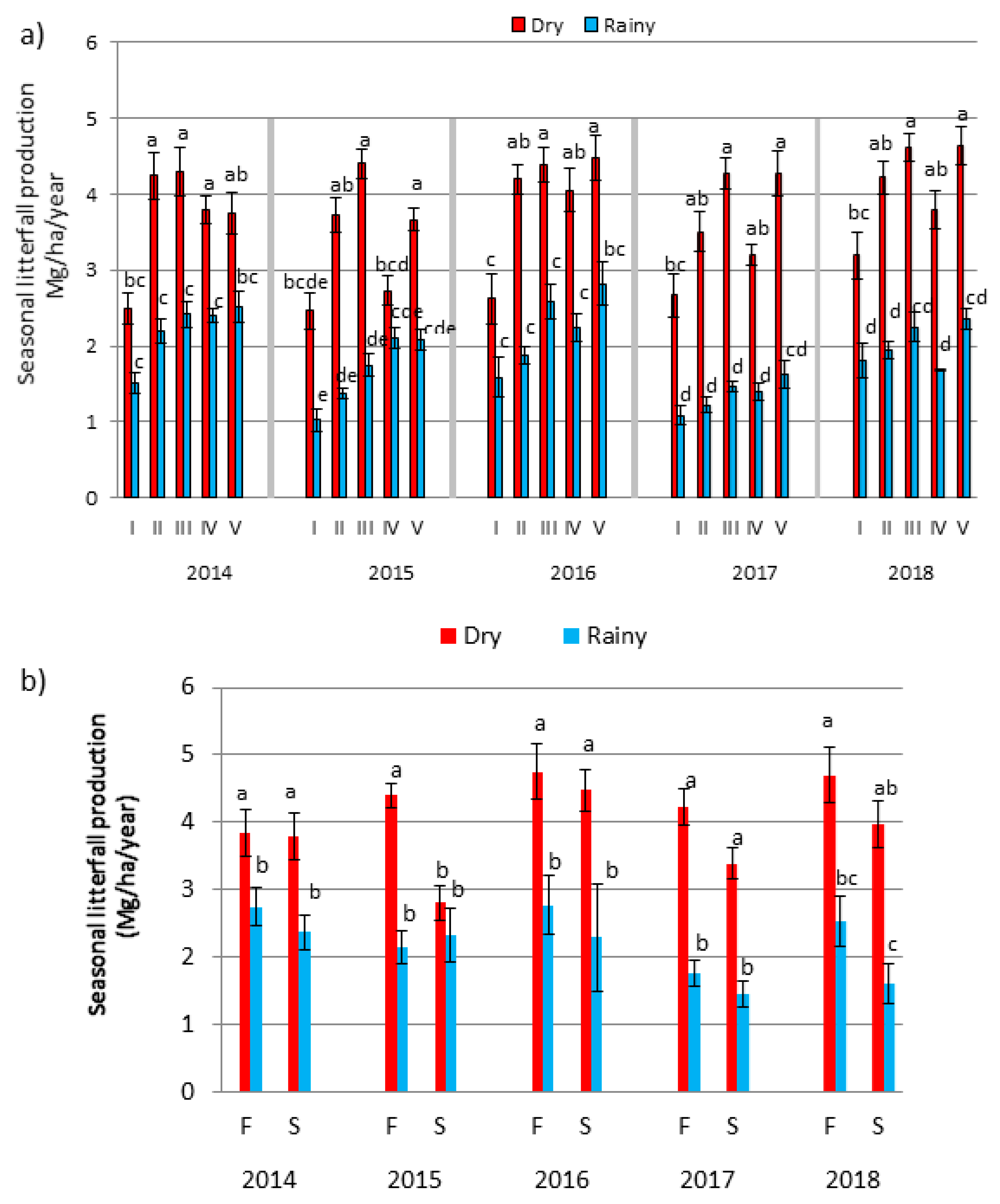
3.1. Total Annual Litterfall Production
3.2. Influence of Successional Age and Topography on Inter-Annual Litterfall Production
3.3. Relationships between Litterfall Production and Vegetation Variables
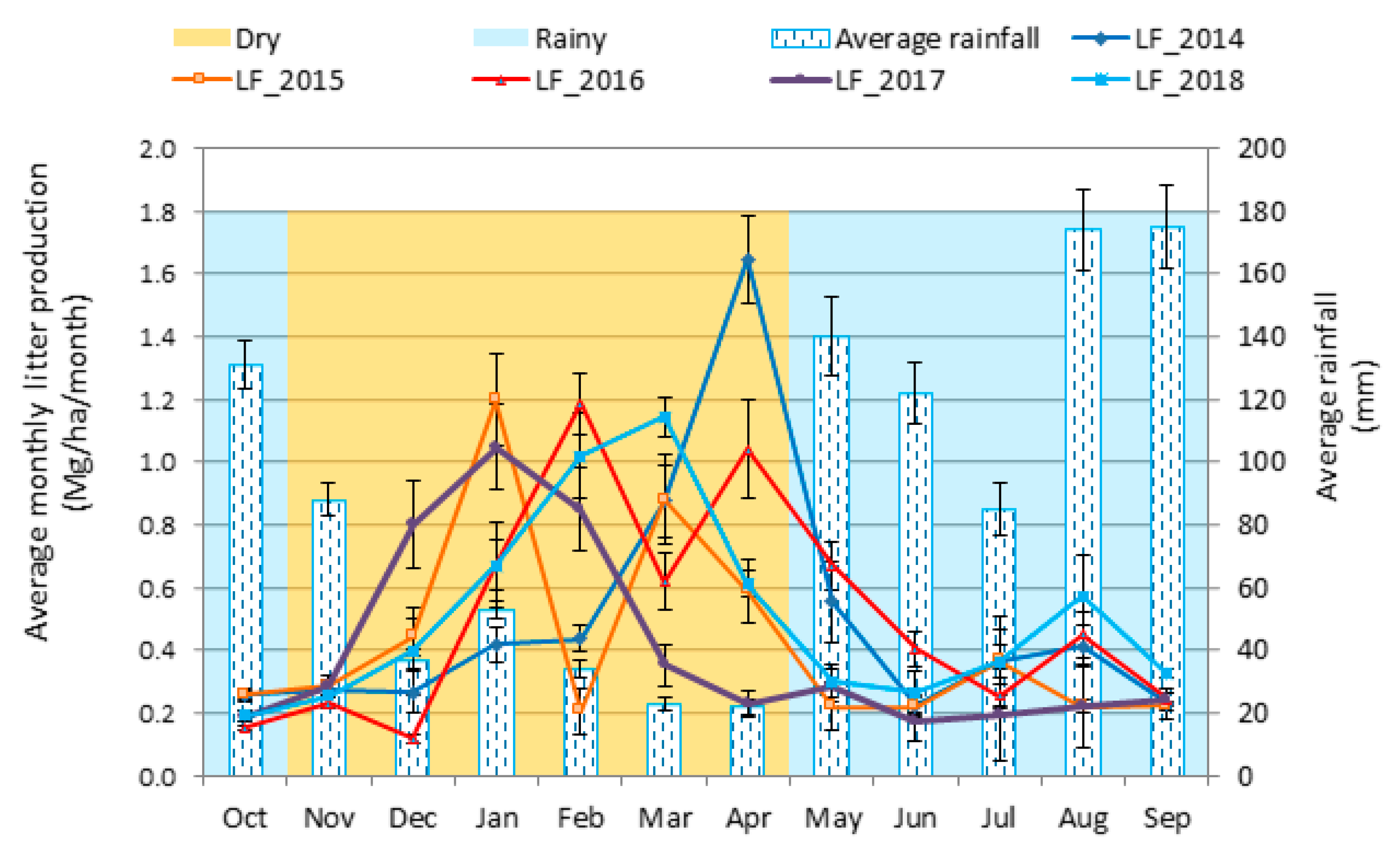
3.4. Seasonal Dynamics of Litterfall Production
3.5. Temporal Variation in Litterfall Production and Its Relationship with Climatic Variables
4. Discussion
4.1. Annual Litterfall Production
4.2. Influence of Successional Age and Vegetation Structure on Litterfall Production
4.3. Influence of Topography and Vegetation Structure on Litterfall Production
4.4. Temporal Variation in Litterfall Production and Its Relationship with Climatic Variables
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster | Age as of 2013 (Years) | Age Class (Years) | Predominant Topographic Position | Average Slope (%) |
|---|---|---|---|---|
| 21 | 3 | I (3–5) | Flat | 2.01 |
| 22 | 4 | I (3–5) | Flat | 1.22 |
| 23 | 3.5 | I (3–5) | Flat | 1.52 |
| 25 | 10 | II (10–17) | Flat | 0 |
| 26 | 17 | II (10–17) | Flat | 0 |
| 28 | 16 | II (10–17) | Flat | 5.01 |
| 29 | 18 | III (18–25) | Flat | 6.63 |
| 30 | 20 | III (18–25) | Flat | 0 |
| 31 | 24 | III (18–25) | Flat | 5.89 |
| 5 | 60 | IV (60–79) | Slope * | 13.41 |
| 7 | 65 | IV (60–79) | Slope * | 14.55 |
| 13 | 70 | IV (60–79) | Flat * | 3.42 |
| 2 | 100 | V (≥80) | Slope * | 15.1 |
| 8 | 80 | V (≥80) | Flat * | 5.49 |
| 20 | 100 | V (≥80) | Flat * | 3.00 |
Appendix B
| No. | Species Name and Authority | Family |
|---|---|---|
| 1 | Acacia dolichostachya S.F. Blake | Fabaceae |
| 2 | Acacia gaumeri S.F. Blake | Fabaceae |
| 3 | Acacia pennatula (Schltdl. & Cham.) Benth. | Fabaceae |
| 4 | Albizia tomentosa (Micheli) Standl. | Fabaceae |
| 5 | Alvaradoa amorphoides Liebm. | Picramniaceae |
| 6 | Amphilophium paniculatum var molle (Schltdl. & Cham.) Standl. | Bignoniaceae |
| 7 | Apoplanesia paniculata C. Presl | Fabaceae |
| 8 | Ardisia escallonioides Schltdl. & Cham. | Primulaceae |
| 9 | Arrabidaea floribunda (Kunth) Loes. | Bignoniaceae |
| 10 | Asemnantha pubescens Hook. f. | Rubiaceae |
| 11 | Attilaea abalak E. Martínez & Ramos | Anacardiaceae |
| 12 | Bauhinia divaricata L. | Fabaceae |
| 13 | Bauhinia ungulata L. | Fabaceae |
| 14 | Bonelia macrocarpa Cav. | Primulaceae |
| 15 | Bourreria pulchra (Millsp.) Millsp. | Ehretiaceae |
| 16 | Bourreria oxyphylla Standl. | Ehretiaceae |
| 17 | Bunchosia glandulosa (Cav.) DC. | Malpighiaceae |
| 18 | Bunchosia swartziana Griseb. | Malpighiaceae |
| 19 | Bursera simaruba (L.) Sarg. | Burseraceae |
| 20 | Caesalpinia gaumeri Greenm. | Fabaceae |
| 21 | Caesalpinia mollis (Kunth) Spreng. | Fabaceae |
| 22 | Caesalpinia yucatanensis Greenm. | Fabaceae |
| 23 | Callicarpa acuminata Kunth. | Lamiaceae |
| 24 | Calyptranthes pallens Griseb. | Myrtaceae |
| 25 | Cardispermum halicacabum L. | Sapindaceae |
| 26 | Casearia emarginata C. Wright ex Griseb. | Salicaceae |
| 27 | Ceiba aesculifolia (Kunth) Britten & Baker f. | Malvaceae |
| 28 | Ceiba pentandra (L.) Gaertn. | Malvaceae |
| 29 | Ceiba schotti Britten & Baker f. | Malvaceae |
| 30 | Celtis iguanaea (Jacq.) Sarg. | Cannabaceae |
| 31 | Ceratophytum tetragonolobum (Jacq.) Sprague & Sandwith | Bignoniaceae |
| 32 | Chiococca alba (L.) Hitchc. | Rubiaceae |
| 33 | Chloroleucon mangense (Jacq.) Britton & Rose | Fabaceae |
| 34 | Chlorophora tinctoria (L.) Gaudich. | Moraceae |
| 35 | Cissus gossypifolia Standl. | Vitaceae |
| 36 | Cnidoscolus aconitifolius (Mill.) I.M. Johnst. | Euphorbiaceae |
| 37 | Coccoloba acapulcensis Standl. | Polygonaceae |
| 38 | Coccoloba diversifolia Jacq. | Polygonaceae |
| 39 | Coccoloba spicata Lundell | Polygonaceae |
| 40 | Cochlospermum vitifolium (Willd.) Spreng. | Bixaceae |
| 41 | Colubrina elliptica (Sw.) Brizicky & W.L. Stern | Rhamnaceae |
| 42 | Colubrina greggii S. Watson | Rhamnaceae |
| 43 | Cordia alliadora (Ruiz & Pav.) Oken | Boraginaceae |
| 44 | Cordia gerascanthus L. | Boraginaceae |
| 45 | Cosmocalyx spectabilis Standl. | Rubiaceae |
| 46 | Croton glabellus L. | Euphorbiaceae |
| 47 | Croton reflexifolius Kunth | Euphorbiaceae |
| 48 | Cydista diversifolia (Kunth) Miers | Bignoniaceae |
| 49 | Cydista heterophylla Seibert | Bignoniaceae |
| 50 | Cydista potosina (K. Schum. & Loes.) Loes. | Bignoniaceae |
| 51 | Cynanchum schlechtendalii (Decne.) Standl. & Steyerm. | Apocynaceae |
| 52 | Diospyros anisandra S.F. Blake | Ebenaceae |
| 53 | Diospyros cuneata Standl. | Ebenaceae |
| 54 | Diospyros salicifolia Humb. & Bonpl. ex Willd. | Ebenaceae |
| 55 | Diospyros verae-crucis (Standl.) Standl. | Ebenaceae |
| 56 | Diospyros yatesiana Standl. ex Lundell | Ebenaceae |
| 57 | Diphysa carthagenensis Jacq. | Fabaceae |
| 58 | Ehretia tinifolia L. | Ehretiaceae |
| 59 | Enterolobium cyclocarpum (Jacq.) Griseb. | Fabaceae |
| 60 | Erythrina standleyana Krukoff | Fabaceae |
| 61 | Erythroxylum rotundifolium Lunan | Erythroxylaceae |
| 62 | Eugenia axillaris (Sw.) Willd. | Myrtaceae |
| 63 | Eugenia buxifolia (Sw.) Willd. | Myrtaceae |
| 64 | Exostema caribaeum (Jacq.) Roem. & Schult. | Rubiaceae |
| 65 | Exostema mexicanum A. Gray | Rubiaceae |
| 66 | Forchhammeria trifoliata Radlk. | Resedaceae |
| 67 | Gliricidia sepium Kunth ex Steud. | Fabaceae |
| 68 | Guettarda combsii Urb. | Rubiaceae |
| 69 | Guettarda elliptica Sw. | Rubiaceae |
| 70 | Guettarda gaumeri Standl. | Rubiaceae |
| 71 | Gymnopodium floribundum Rolfe | Polygonaceae |
| 72 | Hampea trilobata Standl. | Malvaceae |
| 73 | Helicteres baruensis Jacq. | Malvaceae |
| 74 | Heliocarpus donnellsmithii Rose | Malvaceae |
| 75 | Heteropterys brachiata (L.) DC. | Malpighiaceae |
| 76 | Heteropterys laurifolia (L.) A. Juss. | Malpighiaceae |
| 77 | Hippocratea celastroides Kunth | Celastraceae |
| 78 | Hippocratea excelsa Kunth | Celastraceae |
| 79 | Hiraea reclinata Jacq. | Malpighiaceae |
| 80 | Jacquinia macrocarpa Cav. | Primulaceae |
| 81 | Karwinskia humboldtiana (Schult.) Zucc. | Rhamnaceae |
| 82 | Krugiodendron ferreum (Vahl) Urb. | Rhamnaceae |
| 83 | Laetia thamnia L. | Salicaceae |
| 84 | Leucaena leucocephala (Lam.) de Wit | Fabaceae |
| 85 | Lonchocarpus sp Kunth | Fabaceae |
| 86 | Lonchocarpus hondurensis Benth. | Fabaceae |
| 87 | Lonchocarpus longistylus Pittier | Fabaceae |
| 88 | Lonchocarpus rugosus Benth. | Fabaceae |
| 89 | Lonchocarpus xuul Lundell | Fabaceae |
| 90 | Lonchocarpus yucatanensis Pittier | Fabaceae |
| 91 | Luehea candida (DC.) Mart. | Malvaceae |
| 92 | Luehea speciosa Willd. | Malvaceae |
| 93 | Lysiloma latisiliquum (L.) Benth. | Fabaceae |
| 94 | Macfadyena unguis-cati (L.) A.H. Gentry | Bignoniaceae |
| 95 | Machaonia lindeniana Baill. | Rubiaceae |
| 96 | Malpighia glabra L. | Malpighiaceae |
| 97 | Manihot aesculifolia (Kunth) Pohl | Euphorbiaceae |
| 98 | Mansoa verrucifera (Schltdl.) A.H. Gentry | Bignoniaceae |
| 99 | Melicoccus oliviformis Kunth | Sapindaceae |
| 100 | Mimosa bahamensis Benth. | Fabaceae |
| 101 | Montanoa atriplicifolia (Pers.) Sch. Bip. | Asteraceae |
| 102 | Neea psychotrioides Donn. Sm. | Nyctaginaceae |
| 103 | Neea sp. Ruiz & Pav. | Nyctaginaceae |
| 104 | Neomillspaughia emarginata (H. Gross) S.F. Blake | Polygonaceae |
| 105 | Parathesis cubana (A. DC.) Molinet & M. Gómez | Primulaceae |
| 106 | Parmentiera millspaughiana L.O. Williams | Bignoniaceae |
| 107 | Phyllanthus acuminatus Vahl | Phyllanthaceae |
| 108 | Piscidia piscipula (L.) Sarg. | Fabaceae |
| 109 | Pisonia aculeata L. | Nyctaginaceae |
| 110 | Pithecoctenium crucigerum (L.) A.H. Gentry | Bignoniaceae |
| 111 | Platymiscium yucatanum Standl. | Fabaceae |
| 112 | Plumeria obtusa L. | Apocynaceae |
| 113 | Psidium sartorianum (O. Berg) Nied. | Myrtaceae |
| 114 | Psychotria microdon (DC.) Urb. | Rubiaceae |
| 115 | Randia longiloba Hemsl. | Rubiaceae |
| 116 | Randia obcordata S. Watson | Rubiaceae |
| 117 | Randia truncata Greenm. & C.H. Thomps. | Rubiaceae |
| 118 | Sabal japa C. Wright ex Becc. | Arecaceae |
| 119 | Sabal yapa C. Wright ex Becc. | Arecaceae |
| 120 | Samyda yucatanensis Standl. | Salicaceae |
| 121 | Schoepfia schreberi J.F. Gmel. | Schoepfiaceae |
| 122 | Senna atomaria (L.) H.S. Irwin & Barneby | Fabaceae |
| 123 | Senna racemosa (Mill.) H.S. Irwin & Barneby | Fabaceae |
| 124 | Senna villosa (Mill.) H.S. Irwin & Barneby | Fabaceae |
| 125 | Sideroxylon obtusifolium (Humb. ex Roem. & Schult.) T.D. Penn. | Sapotaceae |
| 126 | Solanum erianthum D. Don | Solanaceae |
| 127 | Spondias mombin L. | Anacardiaceae |
| 128 | Tabebuia chrysantha (Jacq.) G. Nicholson | Bignoniaceae |
| 129 | Talisia olivaeformis (Kunth) Radlk. | Sapindaceae |
| 130 | Thevetia gaumeri Hemsl. | Apocynaceae |
| 131 | Thouinia paucidentata Radlk. | Sapindaceae |
| 132 | Trichilia glabra L. | Meliaceae |
| 133 | Vitex gaumeri Greenm. | Lamiaceae |
| 134 | Ximenia americana L. | Ximeniaceae |
| 135 | Zanthoxylum caribaeum Lam. | Rutaceae |
| 136 | Zapoteca Formosa (Kunth) H.M. Hern. | Fabaceae |
Appendix C
| Location | Forest Type | Production (Mg/ha/yr) | Source |
|---|---|---|---|
| Mexico | Subdeciduous | 4.94–6.17 | This study |
| Mexico | Semi-evergreen | 5.20–7.10 | Aryal et al. [16] |
| Mexico | Semi-evergreen and subdeciduous | 3.80–6.80 | Lawrence [21] |
| India | Evergreen and deciduous | 5.76–8.65 | Sundarapandian and Swamy [25] |
| Mexico | Deciduous | 3.98–6.58 | Martínez-Yrízar and Sarukhán [27] |
| Mexico | Semi-evergreen | 5.00–7.70 | Whigham et al. [28] |
| Brazil | Deciduous | 4.00–4.50 | Souza et al. [81] |
References
- Negash, M.; Starr, M. Litterfall production and associated carbon and nitrogen fluxes of seven woody species grown in indigenous agroforestry systems in the south-eastern Rift Valley escarpment of Ethiopia. Nutr. Cycl. Agroecosyst. 2013, 97, 29–41. [Google Scholar] [CrossRef]
- Facelli, J.M.; Pickett, S.T. Plant litter: Its dynamics and effects on plant community. Bot. Rev. 2008, 57, 1–32. [Google Scholar] [CrossRef]
- Dent, D.H.; Bagchi, R.; Robinson, D.; Majalap-Lee, N.; Burslem, D.F.R. Nutrient fluxes via litterfall and leaf litter decomposition vary across a gradient of soil nutrient supply in a lowland tropical rain forest. Plant Soil 2006, 288, 197–215. [Google Scholar] [CrossRef]
- Takyu, M.; Aiba, S.I.; Kitayama, K. Changes in biomass, productivity and decomposition along topographical gradients under different geological conditions in tropical lower montane forests on Mount Kinabalu, Borneo. Oecologia 2003, 134, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Yrízar, A.; Maass, M.; Pérez-Jiménez, L.A.; Sarukhán, J. Net primary productivity of a tropical deciduous forest ecosystem in western Mexico. J. Trop. Ecol. 1996, 12, 169–175. [Google Scholar] [CrossRef]
- González, R.H.; Cantú, I.; Ramírez, G.R.; Gómez, M.V.; Domínguez, T.G.; Bravo, J.; Maiti, R.K. Spatial and seasonal litterfall deposition pattern in the Tamaulipan thorscrub, Northeastern Mexico. J. Environ. 2008, 1, 177–181. [Google Scholar]
- Bosco, J.I.; Blanco, J.A.; Castillo, F.J. Gestión forestal y ciclos de nutrientes en el marco del cambio global. In Ecología de Bosque Mediterráneo en un Mundo Cambiante; Valladares, F., Ed.; Ministerio del Medio Ambiente: Madrid, Spain, 2004; pp. 479–506. [Google Scholar] [CrossRef] [Green Version]
- Prause, J.; De Caram, A.G.; Angeloni, P.N. Variación mensual en el aporte de cuatro especies forestales nativas del Parque Chaqueño Humado (Argentina). Quebracho 2003, 10, 39–45. [Google Scholar]
- Melillo, J.M.; McGuire, A.D.; Kicklighter, D.W.; Moore, B.; Vorosmarty, C.J.; Schloss, A.L. Global climate change and terrestrial net primary production. Nature 1993, 363, 234–240. [Google Scholar] [CrossRef]
- De Jong, B.H.J. Spatial distribution of biomass and links to reported disturbances in tropical lowland forests of southern Mexico. Carbon Manag. 2013, 4, 601–615. [Google Scholar] [CrossRef]
- Scheer, M.B.; Gatti, G.; Wisniewski, C. Nutrient fluxes in litterfall of a secondary successional alluvial rain forest in Southern Brazil. Rev. Biol. Trop. 2011, 59, 1869–1882. [Google Scholar]
- Thuille, A.; Schulze, E.D. Carbon dynamics in successional and afforested spruce stands in Thuringia and the Alps. Glob. Chang. Biol. 2006, 12, 325–342. [Google Scholar] [CrossRef]
- Sánchez-Azofeifa, A.; Powers, J.; Fernandes, G.; Quesada, M. Tropical dry forest in the Americas: The Tropi-Dry Endeavor. In Tropical Dry Forests in the Americas: Ecology, Conservation and Management; Sánchez-Azofeifa, A., Power, J., Fernandes, G., Quesada, M., Eds.; Routledge: New York, NY, USA, 2013; pp. 20–34. [Google Scholar]
- Banda-R, K.; Delgado, S.A.; Dexter, K.G.; Linares, P.R.; Oliveira, F.A.; Prado, D.; Pullan, M.; Quintana, C.; Riina, R.; Rodríguez, G.M.; et al. Plant diversity patterns in neotropical dry forests and their conservation implications. Science 2016, 353, 1383–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huechacona-Ruíz, A.H. Dinámica de la Producción de Hojarasca y el Índice de Área Foliar en un Bosque Tropical Seco en Yucatán. Master’s Thesis, Centro de Investigación Científica de Yucatán, Mérida, Mexico, 2016. [Google Scholar] [CrossRef]
- Aryal, D.B.; de Jong, B.H.J.; Ochoa-Gaona, S.; Mendoza-Vega, J.; Esparza-Olguin, L. Successional and seasonal variation in litterfall and associated nutrient transfer in semi-evergreen tropical forests of SE Mexico. Nutr. Cycl. Agroecosyst. 2015, 103, 45–60. [Google Scholar] [CrossRef]
- Valentini, C.M.A.; Sanches, L.; de Paula, S.R.; Vourlitis, G.L.; Nogueira, J.D.; Pinto, O.B.P.; Lobo, F.D. Soil respiration and aboveground litter dynamics of a tropical transitional forest in northwest Mato Grosso, Brazil. J. Geophys. Biogeosci. Res. 2008, 113, 11. [Google Scholar] [CrossRef]
- Martins, S.V.; Rodrigues, R.R. Produção de serapilheira em clareiras de uma floresta estacional semidecidual no municipio de Campinas, SP. Rev. Bras. Bot. 1999, 22, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yuan, W.; Dong, W.; Liu, S. Seasonal patterns of litterfall in forest ecosystem worldwide. Ecol. Complex 2014, 20, 240–247. [Google Scholar] [CrossRef]
- Cuba, N.; Rogan, J.; Christman, Z.; Williams, C.; Schneider, L.; Lawrence, D.; Millones, M. Modelling dry season deciduousness in Mexican Yucatán forest using MODIS EVI data (2000–2011). GISci. Remote Sens. 2013, 50, 26–49. [Google Scholar] [CrossRef]
- Lawrence, D. Regional-scale variation in litter production and seasonality in tropical dry forests of Southern Mexico. Biotropica 2005, 37, 561–570. [Google Scholar] [CrossRef]
- Kelly, D.L.; Tanner, E.V.; Kapos, V.; Dickinson, T.A.; Goodreiend, G.A.; Fiiarbairn, P.I. Jamaican limestone forests: Floristries, structure and environment of three examples along a rainfall gradient. J. Trop. Ecol. 1988, 24, 121–156. [Google Scholar] [CrossRef]
- Murphy, P.G.; Lugo, A. Ecology of tropical dry Forest. Annu. Rev. Ecol. Syst. 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Parsons, S.A.; Congdon, R.A.; Shoo, L.P.; Valdez, V.; Williams, S.E. Spatial variability in litterfall, litter standing crop and litter quality in a tropical rain forest region. Biotropica 2014, 46, 378–386. [Google Scholar] [CrossRef]
- Sundarapandian, S.M.; Swamy, P.S. Litter production and leaf litter decomposition of selected tree species in tropical forests at Kodayar in the Western Ghats, India. For. Ecol. Manag. 1999, 123, 231–244. [Google Scholar] [CrossRef]
- Sánchez-Silva, S.; De Jong, B.H.J.; Aryal, D.R.; Huerta-Lwanga, E.; Mendoza-Vega, J. Trends in leaf traits, litter dynamics and associated nutrient cycling along a secondary successional chronosequence of semi-evergreen tropical forest in South-Eastern Mexico. J. Trop. Ecol. 2018, 34, 364–377. [Google Scholar] [CrossRef]
- Martínez-Yrízar, A.; Sarukhán, J. Litterfall patterns in a tropical deciduous forest in Mexico over a five-year period. J. Trop. Ecol. 1990, 6, 137–154. [Google Scholar] [CrossRef]
- Whigham, D.F.; Zugasty, P.; Cabrera, E.; O’neill, J.; Ley, E. The effecto of annual variation in precipitation on growth and litter production in a tropical dry forest in the Yucatan of Mexico. J. Trop. Ecol. 1990, 32, 23–34. [Google Scholar]
- Tang, J.; Cao, M.; Zhang, J.; Li, M. Litterfall production, decomposition and nutrient use efficiency varies with tropical forest types in Xishuangbanna, SW China: A 10-year study. Plant Soil 2010, 335, 271–288. [Google Scholar] [CrossRef]
- Wagner, F.H.; Hérault, B.; Bonal, D.; Stahl, C.; Anderson, L.O.; Baker, T.R.; Becker, G.S.; Beeckman, H.; Souza, D.B.; Botosso, P.C.; et al. Climate seasonality limits leaf carbon assimilation and wood productivity in tropical forests. Biogeosciences 2016, 13, 2537–2562. [Google Scholar] [CrossRef] [Green Version]
- Barlow, J.; Gardner, T.A.; Ferreira, L.V.; Pérez, C.A. Litter fall and decomposition in primary, secondary and plantation forest in the Brazilian Amazon. For. Ecol. Manag. 2007, 247, 91–97. [Google Scholar] [CrossRef]
- Huang, Y.; Ma, Y.; Zhao, K.; Niklaus, P.A.; Schmid, B.; He, J. Positive effects of tree species diversity on litterfall quantity and quality along a secondary successional chronosequence in a subtropical forest. Plant Ecol. 2017, 10, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Zalamea, M.; González, G. Leaf fall phenology in a subtropical we forest in Puerto Rico: From species to community patterns. Biotropica 2008, 40, 295–304. [Google Scholar] [CrossRef]
- Holbrook, N.M.; Whitbeck, J.L.; Mooney, H.A. Drought responses of Neotropical dry forest trees. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 243–276. [Google Scholar] [CrossRef]
- Gaspar-Santos, E.; Espinoza, M.G.; Marcial, N.R.; Álvarez, S.J.D. Acumulación y descomposición de hojarasca en bosques secundarios del sur de la Sierra Madre de Chiapas, México. Bosque 2015, 36, 467–480. [Google Scholar] [CrossRef] [Green Version]
- Celentano, D.; Zahawi, R.A.; Finegan, B.; Ostertag, R.; Cole, R.J.; Holl, K.D. Litterfall dynamics under different tropical forest restoration strategies in Costa Rica. Biotropica 2011, 43, 279–287. [Google Scholar] [CrossRef]
- Chave, J.; Navarrete, D.; Almeida, S.; Álvarez, E.; Aragão, L.E.; Bonal, D.; Châtelet, P.; Silva-Espejo, J.; Goret, J.Y.; von Hildebrand, P.; et al. Regional and temporal patterns of litterfall in tropical South America. Biogeosci. Discuss 2009, 6, 7565–7597. [Google Scholar] [CrossRef]
- Norden, N.; Angarita, H.A.; Bongers, F.; Martínez, M.; La Cerda, I.G.; Van Breugel, M.; Lebrija-Trejos, E.; Meave, J.A.; Vandermeer, J.; Williamson, G.B.; et al. Successional dynamics in Neotropical forests are as uncertain as they are predictable. Proc. Natl. Acad. Sci. USA 2015, 112, 803–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, Y.R.F.; Luz, G.R.; Souza, S.R.; Silva, D.; Veloso, M.D.M.; Espírito-Santo, M.M.; Santos, R.M. Floristic, structural, and functional group variations in tree assemblages in a Brazilian tropical dry Forest: Effects of successional stage and soil properties. In Tropical Dry Forests in the Americas: Ecology, Conservation, and Management; Sánchez-Azofeifa, A., Powers, J.S., Fernandes, G.W., Quesada, M., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 329–353. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Pérez, E.A.; Meave, J.A.; Poorter, L.; Bongers, F. Environmental changes during secondary succession in a tropical dry forest in Mexico. J. Trop. Ecol. 2011, 27, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Chazdon, R.L.; Letcher, S.G.; Van Breugek, M.; Martínez-Ramos, M. Rates of change in tree communities of secondary neotropical forests following major disturbances. Philos. Trans. R. Soc. Lond. Ser. B 2007, 362, 273–289. [Google Scholar] [CrossRef]
- Lohbeck, M.; Lebrija-Trejos, E.; Martinez-Ramos, M.; Meave, J.A.; Poorter, L.; Bongers, F. Functional trait strategies of trees in dry and wet tropical forests are similar but differ in their consequences for succession. PLoS ONE 2015, 10, e0123741. [Google Scholar] [CrossRef] [Green Version]
- Schilling, J.S.; Ayres, A.; Kaffenberger, J.T.; Powers, J.S. Initial white rot type dominance of wood decomposition and its functional consequences in a regenerating tropical dry forest. Soil Biol. Biochem. 2016, 88, 58–68. [Google Scholar] [CrossRef]
- Lasky, J.R.; Uriarte, M.; Boukili, V.K.; Chazdon, R.L. Trait-mediated assembly processes predict successional changes in community diversity of tropical forests. Proc. Natl. Acad. Sci. USA 2014, 111, 5616–5621. [Google Scholar] [CrossRef] [Green Version]
- Becknell, J.M.; Powers, J.S. Stand age and soils as drivers of plant functional traits and aboveground biomass in secondary tropical dry forest. Can. J. For. Res. 2014, 44, 604–613. [Google Scholar] [CrossRef]
- Quesada, M.; Álvarez, M.; Avila, L.; Castillo, A.; Lopezaraiza, M.; Martén, V.; Guerrero, V.R.; Sáyago, R.G.; Sánchez, G.; Contreras, J.M.; et al. Tropical Dry Forest Ecological Succession in México: Synthesis of a Long-Term Study. In Tropical Dry Forest in the Americas; Sánchez-Azofeifa, A., Powers, J., Fernandes, G., Quesada, M., Eds.; Island Press: Washington, DC, USA, 2009; pp. 17–53. [Google Scholar]
- Guariguata, M.R.; Osterbag, R. Neotropical secondary forest succession: Changes in structural and functional characteristics. For. Ecol. Manag. 2001, 148, 185–206. [Google Scholar] [CrossRef]
- Buzzard, V.; Hulshof, C.M.; Birt, T.; Violle, C.; Enquist, B.J. Data from: Re-growing a tropical dry forest: Functional plant trait composition and community assembly during succession provisional. Dryad Digit. Repos. 2016. [Google Scholar] [CrossRef]
- Lohbeck, M.; Poorter, L.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Paz, H.; Pérez-García, E.A.; Romero, I.E.; Tauro, A.; Bongers, F. Successional changes in functional composition contrast for dry and wet tropical forest. Ecology 2013, 94, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Sanaphre-Villanueva, L.; Dupuy, J.M.; Andrade, J.L.; Reyes, C.; Jackson, P.C.; Paz, H. Patterns of plant functional variation and specialization along secondary succession and topography in a tropical dry forest. Environ. Res. Lett. 2017, 12, 1–10. [Google Scholar] [CrossRef]
- Pineda-García, F.; Paz, H.; Meinzer, F.C. Drought resistance in early and late secondary successional species from a tropical dry forest: The interplay between xylem resistance to embolism, sapwood water storage and leaf shedding. Plant Cell. Environ. 2013, 36, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Lebrija-Trejos, E.; Meave, J.A.; Poorter, L.; Pérez, E.A.; Bongers, F. Pathways, mechanisms and predictability of vegetation change during tropical dry forest succession. Annu. Rev. Ecol. Syst. 2010, 12, 267–275. [Google Scholar] [CrossRef]
- Méndez-Alonzo, R.; Pineda, F.; Paz, H.; Rosell, J.A.; Olson, M.E. Leaf phenology is associated with soil water availability and xylem traits in a tropical dry forest. Trees 2013, 27, 745–754. [Google Scholar] [CrossRef]
- Gallardo-Cruz, J.A.; Pérez-García, E.A.; Meave, J.A. Diversity and vegetation structure as influenced by slope aspect and altitude in a seasonally dry tropical landscape. Landsc. Ecol. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Pierce, K., Jr.; Lookingbill, T.; Urban, D. A simple method for estimating potential relative radiation (PRR) forlandscape-scale vegetation analysis. Landsc. Ecol. 2005, 20, 137–147. [Google Scholar] [CrossRef]
- Suzaki, T.; Kume, A.; Ino, Y. Effects of slope and canopy trees on light conditions and biomass of dwarf bamboo under acoppice canopy. J. For. Res. 2005, 10, 151–156. [Google Scholar] [CrossRef]
- Becker, P.; Rabenold, P.E.; Idol, J.R.; Smith, A.P. Water potential gradients for gaps and slopes in a Panamanian tropical moist forest’s dry season. J. Trop. Ecol. 1988, 4, 173–184. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L.; Paz, H.; Sack, L.; Bongers, F. Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits. Plant Cell. Environ. 2010, 34, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Daws, M.I.; Mullins, C.E.; Burslem, D.F.; Paton, S.R.; Dalling, J.W. Topographic position affects the water regime in a semideciduous tropical forest in Panama. Plant Soil 2002, 238, 79–89. [Google Scholar] [CrossRef]
- Borchert, R. Soil and Stem Water Storage Determine Phenology and Distribution of Tropical Dry Forest Trees. Ecology 1994, 75, 1437. [Google Scholar] [CrossRef]
- Orellana, R.; Islebe, G.; Espadas, C. Presente, pasado y futuro de los climas de la Península de Yucatán. In Naturaleza y Sociedad en el Área Maya: Pasado, Presente y Futuro; Colunga, M.P., Larqué, A., Eds.; Centro de Investigación Científica de Yucatán: Mérida, Mexico, 2003; pp. 37–52. [Google Scholar] [CrossRef] [Green Version]
- Bautista-Zuñiga, F.; Batllori-Sampedro, E.; Ortiz-Pérez, M.A.; Palacio-Aponte, G.; Castillo-González, M. Geoformas, agua y suelo en la Península de Yucatán. In Naturaleza y Sociedad en el Área Maya: Pasado, Presente y Futuro; Colunga, M.G., Patricia, S., Larqué, A.S., Eds.; Academia Mexicana de Ciencias, Centro de Investigación Científica de Yucatán: Mérida, Mexico, 2003; pp. 21–36. [Google Scholar] [CrossRef]
- Dupuy, J.M.; Hernández-Stefanoni, J.L.; Hernández, R.A.; Tetetla-Rangel, E.; López, J.O.; Martínez, E.; Leyequién-Abarca, E.; Tun-Dzul, F.; May Pat, F. Patterns and correlates of tropical dry forest structure and composition in a highly replicated chronosequence in Yucatan, Mexico. Biotropica 2012, 44, 151–162. [Google Scholar] [CrossRef]
- Hernández-Stefanoni, J.L.S.; Dupuy, J.M.; Tun-Dzul, F.; May-Pat, F. Influence of landscape structure and stand age on species density and biomass of a tropical dry forest across spatial scales. Landsc. Ecol. 2011, 26, 355–370. [Google Scholar] [CrossRef]
- Comisión Nacional Forestal (CONAFOR). Manual y Procedimiento Para el Muestreo de Campo; Comisión Nacional Forestal: Jalisco, Mexico, 2013. [Google Scholar]
- Aceñolaza, P.G.; Zamboni, L.P.; Gallardo, J.F. Aporte de hojarasca en bosques del predelta del río Paraná (Argentina). Bosque 2009, 30, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Honorio, E.N.; Baker, T.R. Manual Para el Monitoreo del Ciclo del Carbono en Bosques Amazónicos; Instituto de Investigaciones de la Amazonia Peruana, Universidad de Leeds: Lima, Peru, 2010. [Google Scholar] [CrossRef]
- Jones, H.G. Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology, 2nd ed.; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Ramírez, G.; Dupuy, J.M.; Solorio, F.J.; Ramírez, L. Ecuaciones Alométricas de Biomasa Epigea en Una Selva Mediana Subcaducifolia de la Península de Yucatán. Ph.D. Thesis, Centro de Investigación Científica de Yucatán, Mérida, Mexico, 2019. [Google Scholar]
- Chave, J.; Andalo, S.; Brown, M.A.; Cairns, J.Q.; Chambers, D.; Eamus, H.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stock and balance in tropical forests. Oecología 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Duno, R.; Carnevali, G.; Ramírez, I.M.; Tapia, J.L.M.; Lorena, L.C.; Hernández, S.A.; Embray, T. Flora Ilustrada de la Península de Yucatán: Listado Florístico; Centro de Investigación Científica de Yucatán: Mérida, Mexico, 2010. [Google Scholar] [CrossRef]
- Pennington, T.D.; Sarukhán, J. Árboles Tropicales de México. Manual Para la Identificación de las Principales Especies, 3rd ed.; Ediciones Científicas Universitarias, UNAM: Mexico City, Mexico, 2005. [Google Scholar] [CrossRef]
- Anselin, L. Local Indicators of Spatial Association-ISA. Geogr. Anal. 2010, 27, 93–115. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2020. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2020. Available online: https://www.qgis.org/en/site/ (accessed on 24 January 2020).
- SPSS Inc. SPSS Statistics Para Windows, versión 17.0; SPSS Inc.: Chicago, IL, USA, 2008.
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012; Available online: https://www.R-project.org/ (accessed on 22 June 2018).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Caballero, D.F.F. Selección de Modelos Mediante Criterios de Información en Análisis Factorial. Aspectos Teóricos y Computacionales. Ph.D. Thesis, Universidad de Granada, Granada, Spain, 2011. [Google Scholar]
- Ichikawa, M. Empirical assessments of AIC procedure for model selection in factor analysis. Behaviormetrika 1988, 24, 499–509. [Google Scholar] [CrossRef]
- Souza, S.R.; Veloso, M.D.M.; Espírito-Santo, M.M.; Silva, J.O.; Sánchez-Azofeifa, A.; Souza e Brito, B.G.; Fernandez, G.W. Litterfall dynamics along a successional gradient in a Brazilian tropical dry forest. For. Ecosyst. 2019, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Guardado, M.A. Estimación del Carbono Retenido en la Biomasa Aérea en Una Selva Tropical de Yucatán, México. Master’s Thesis, Centro de Investigación Científica de Yucatán, Mérida, Mexico, 2015. [Google Scholar]
- Ruiz, J.; Fandino, M.C.; Chazdon, R.L. Vegetation structure, composition, and species richness across a 56-year chronosequence of dry tropical forest on Providencia Island, Colombia. Biotropica 2005, 37, 520–530. [Google Scholar] [CrossRef]
- Ceccon, E.; Sánchez, S.; Campo, J. Tree seedling dynamics in two abandoned tropical dry forest of differing successional status in Yucatan, México: Field experiment whit N and P fertilization. Plant Ecol. 2004, 170, 277–285. [Google Scholar] [CrossRef]
- Feng, C.; Wang, Z.; Ma, Y.; Fu, S.; Chen, H.Y. Increased litterfall contributes to carbon and nitrogen accumulation following cessation of anthropogenic disturbances in degraded forests. For. Ecol. Manag. 2019, 432, 832–839. [Google Scholar] [CrossRef]
- Pineda, F.; Paz, H.; Tinoco, C. Morphological and physiological differentiation of seedlings between dry and wet habitats in a tropical dry forest: Water-use strategies of TDF tree seedlings. Plant Cell. Environ. 2011, 34, 1536–1547. [Google Scholar] [CrossRef] [PubMed]
- Pineda-García, F.; Paz, H.; Meinzer, F.C.; Angeles, G. Exploiting water versus tolerating drought: Water-use strategies of trees in a secondary successional tropical dry forest. Tree Physiol. 2015, 36, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Markesteijn, L.; Poorter, L. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought-and shade-tolerance. Open J. Ecol. 2009, 97, 311–325. [Google Scholar] [CrossRef]
- Méndez, R.; Paz, H.; Zuluaga, R.C.; Rosell, J.A.; Olson, M.E. Coordinated evolution of leaf and stem economics in tropical dry forest trees. Ecology 2012, 93, 2397–2406. [Google Scholar] [CrossRef]
- Gei, M.G.; Rozendaal, D.M.A.; Poorter, L.; Bongers, F.; Sprent, J.; Mira, G.D.; Aide, T.M.; Andrade, J.L.; Balvanera, P.; Becknell, J.M.; et al. Legume Abundance Along Successional And Rainfall Gradients In Neotropical Forests. Nat. Ecol. Evol. 2018, 2, 1104–1111. [Google Scholar] [CrossRef]
- Erickson, H.E.; Helmer, E.H.; Bandeis, T.J.; Lugo, A.E. Control on fallen leaf chemistry and forest floor element masses in native and forest across a tropical island. Ecosphere 2014, 5, 1–28. [Google Scholar] [CrossRef] [Green Version]
- López-Martínez, J.O.; Sanaphre, L.; Dupuy, J.M.; Hernández-Stefanoni, J.L.; Meave, J.A.; Gallardo, J.A. β-Diversity of Functional Groups of Woody Plants in a Tropical Dry Forest in Yucatan. PLoS ONE 2013, 8, e73660. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Stefanoni, J.L.; Dupuy, J.M.; Tun-Dzul, F.; May Pat, F. Influencia de la estructura del paisaje y la edad de sucesión sobre la diversidad de especies y la biomasa aérea a diferentes escalas. In Planeación de Uso del Suelo a Nivel de Paisaje en el Sur de Yucatán; Centro de Investigación Científica de Yucatán: Mérida, Mexico, 2012; pp. 51–64. [Google Scholar] [CrossRef]
- Nafarrate-Hecht, A.C. Estimación Directa e Indirecta del Índice de Área Foliar y su Modelación con Lidar en un Bosque Tropical Seco de Yucatán. Master’s Thesis, Centro de Investigación Científica de Yucatán, Mérida, Mexico, 2017. [Google Scholar] [CrossRef]
- Valdez, M.; Andrade, J.L.; Jackson, P.C.; Rebolledo, M. Phenology of five tree species of a tropical dry forest in Yucatan, México: Effects of environmental and physiological factors. Plant Soil 2010, 329, 155–171. [Google Scholar] [CrossRef]
- Portillo, C.A.; Sánchez-Azofeifa, A. Extent and conservation of tropical dry forests in the Americas. Biol. Conserv. 2009, 143, 144–155. [Google Scholar] [CrossRef]
- Smith, V.C.; Ennos, R. The effects of air flow and stem flexure on the mechanical and hydraulic properties of the stems of sunflowers Helianthus annuus L. J. Exp. Bot. 2003, 845–849. [Google Scholar] [CrossRef] [PubMed]
- Van Gardingen, P.R.; Grace, J. Plants and wind. Bot. Res. 1991, 18, 192–253. [Google Scholar]
- Yazaki, T.; Hirano, T.; Sano, T. Biomass accumulation and net primary production during the early stage of secondary succession after a severe forest disturbance in northern Japan. Forests 2016, 7, 287. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.C.A.; Araújo, V.F.P.; Vasconcellos, A. The effects of rainfall and vegetation on litterfall production in the semiarid region of northeastern Brazil. Braz. J. Biol. 2015, 75, 703–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzini, F.F.; Ranieri, B.D.; Brandão, D.; Fernandes, G.W.; Quesada, M.; Espírito-Santo, M.M.; Jacobi, C.M. Changes in tree phenology along natural regeneration in a seasonally dry tropical forest. Plant Biosyst. 2014, 148, 965–974. [Google Scholar] [CrossRef]
- Fernández, E.; Vázquez, T.; Hidalgo, J.Z.; Centeno, R.R. Actualización de los Escenarios de Cambio Climático para Estudios de Impactos, Vulnerabilidad y Adaptación; Unidad Informática para las Ciencias Atmosféricas y Ambientales, Centro de Ciencias de la Atmósfera, Universidad Nacional Autónoma de México, Instituto Nacional de Ecología y Cambio Climático. 2015. Available online: http://atlasclimático.unam.mx/ACM/ (accessed on 10 September 2020).
- Ellis, E.A.; Romero Montero, A.; Hernández Gómez, I.U. Evaluación y Mapeo de los Determinantes de Deforestación en la Península Yucatán; USAID-TNC-MREDD+; Mexico, 2015. Available online: https://www.researchgate.net/profile/Edward_Ellis/publication/283090392_Evaluacion_y_mapeo_de_los_determinantes_de_la_deforestacion_en_la_Peninsula_Yucatan-USAIDTNCMREDD/links/562a7d7808ae518e347f54dc.pdf (accessed on 15 September 2020). [CrossRef]
- Esparza, O.L.G.; Martínez, E.R. Deforestación en Campeche: Causas y Efectos. Rev. Fomix Campeche 2011, 3, 6–11. [Google Scholar]
- Ramírez, L.C.; Rivera, L.J.A. La ganadería en el contexto de la biodiversidad. In Biodiversidad y Desarrollo Humano en Yucatán; Durán, R., Méndez, M., Eds.; PPF-FMAM; Conabio, Seduma: Mérida, Mexico, 2010. [Google Scholar]
- Sohn, Y.; Moran, E.; Gurri, F. Deforestation in north-central Yucatán (1985–1995): Mapping secundary succession of forest and agricultural land use in Sotuta using the cosine of the angle concept. In Environmental Sciencie and Engineer for 21st Century; National Science Foundation: Alexandria, VA, USA, 1999; Volume 65, pp. 947–958. [Google Scholar]




| Year | Annual Litterfall Production (Mg/ha/yr) | Annual Precipitation (mm) |
|---|---|---|
| 2014 | 5.963 ± 0.955 “ab” | 1148.0 |
| 2015 | 5.071 ± 0.603 “ab” | 1098.8 |
| 2016 | 6.173 ± 0.654 “a” | 885.2 |
| 2017 | 4.941 ± 0.502 “b” | 906.5 |
| 2018 | 6.109 ± 0.508 “a” | 945.2 |
| Successional Age Class | Total Litterfall Production (Mg/ha/yr) |
|---|---|
| I (3–5 years) | 4.070 ± 0.502 “a” |
| II (10–17 years) | 5.674 ± 0.274 “b” |
| III (18–25 years) | 6.460 ± 0.173 “c” |
| IV (60–79 years) | 5.604 ± 0.502 “b” |
| V (≥80 years) | 6.440 ± 0.281 “c” |
| Year | Models | R2aj | AIC | ∆ AIC | ωi |
|---|---|---|---|---|---|
| 2014 | (1) Y = 0.885 × Ht | 0.379 | 13.77 | 2.69 | 0.004 |
| (2) Y = 1.371 × Ht − 0.044 × Be | 0.491 | 11.54 | 0.46 | 0.666 | |
| (3) Y = 0.02 × E − 0.054 × Be + 1.16Ht | 0.529 | 11.08 | 0.00 | 0.003 | |
| 2015 | (1) Y = 1.604 + 0.532 × Ht | 0.424 | −1.156 | 0.642 | 0.079 |
| (2) Y = 0.141 × BAd | 0.464 | −1.424 | 0.374 | 0.255 | |
| (3) Y* = −0.037 + 0.966 × Ht + 0.019 × Bd | 0.500 | −1.798 | 0.000 | 0.666 | |
| 2016 | (1) Y = 1.812 + 0.684 × Ht | 0.624 | −5.179 | 0.000 | 0.079 |
| (2) Y* = 1.437 − 0.04 × BAe + 0.793 × Ht | 0.608 | −3.734 | 1.445 | 0.255 | |
| (3) Y = 1.74 + 0.009 × Age | 0.607 | −3.007 | 2.172 | 0.666 | |
| 2018 | (1) Y = 3.819 + 0.718 × Ht | 0.243 | −2.260 | 4.270 | 0.079 |
| (2) Y = 2.09 + 0.224 × DBH | 0.320 | −3.054 | 1.913 | 0.255 | |
| (3) Y” =1.053 + 0.292 × DBH + 0.01 × BAd | 0.322 | −2.408 | 0.000 | 0.000 | |
| Total | (1) Y = 3.772 + 0.115 × BAd | 0.516 | −2.670 | 4.270 | 0.124 |
| (2) Y = 4.204 + 0.034 × Bd − 0.158 × BAe | 0.608 | −5.027 | 1.920 | 0.452 | |
| (3) Y* = 3.761 + 0.061 × Abd + 0.024 × Bd − 0.141 × BAe | 0.607 | −6.940 | 0.000 | 0.424 |
| Model | R2aj | AIC | ∆ AIC | Ѡi |
|---|---|---|---|---|
| annual | ||||
| (1) Y = 9.328 − 0.003 × P | 0.944 | −18.17 | 33.41 | 0.124 |
| (2) Y* = 5.18 + 0.067 × RH − 0.04 × P | 0.981 | −24.81 | 26.77 | 0.452 |
| (3) Y* = −0.0015 × P + 0.001 × W + 0.00009 × ACTmin | 0.990 | −51.58 | 0.000 | 0.424 |
| seasonal | ||||
| (1) Y = −7.345 + 2.071 × W | 0.629 | 5.758 | 9.744 | 0.009 |
| (2) Y” = 1.88 + 1.87 × V − 0.32 × Tmean | 0.692 | 4.549 | 8.535 | 0.452 |
| (3) Y = 2.16 × W + 0.0019 × ACTmax | 0.874 | −3.986 | 0.000 | 0.105 |
| Year | Model | R2aj | AIC | ∆ AIC | ωi |
|---|---|---|---|---|---|
| 2014 | (1) Y = 4.215 − 0.045 × RH | 0.77 | −37.96 | 0.37 | 0.332 |
| (2) Y = 1.625 − 0.06 × Tmax + 1.63 × DPV | 0.79 | −38.33 | 0.00 | 0.399 | |
| (3) Y*= 0.17 × RH − 0.21 × T + 6.81 × DPV | 0.79 | −37.54 | 0.79 | 0.269 | |
| 2015 | (1) Y”= 0.81 − 0.032 × WG | 0.11 | −30.54 | 0.34 | 0.303 |
| (2) Y”= 0.24 − 0.054 × Tmin + 0.0009 × ACTmax | 0.18 | −30.75 | 0.13 | 0.337 | |
| (3) Y”= −0.02 × WG − 0.05 × Tmin + 0.001 × ACTmax | 0.23 | −30.88 | 0.00 | 0.360 | |
| 2016 | (1) Y= 1.31 − 0.059 × Tmin | 0.45 | −30.83 | 10.83 | 0.003 |
| (2) Y= -0.023 × RH − 0.003 × ACTmin | 0.75 | −39.87 | 1.79 | 0.289 | |
| (3) Y= 0.076 × Tmax − 0.003 × ACTmin | 0.80 | −41.66 | 0.00 | 0.720 | |
| 2017 | (1) Y = 3.95 − 0.131 × Tmean | 0.43 | −38.56 | 4.63 | 0.133 |
| (2) Y = 3.391 − 0.094 × Tmean − 0.035 ×Tmin | 0.76 | −43.05 | 0.14 | 0.433 | |
| (3) Y*= −0.37 + 0.13 × RH − 0.37 × Tmean + 4.08 × DPV | 0.77 | −43.19 | 0.00 | 0.433 | |
| 2018 | (1) Y = 1.606 − 0.014 × RH | 0.45 | −33.60 | 3.12 | 0.049 |
| (2) Y = 1.533 − 0.011 × RH − 0.001 × P | 0.55 | −35.36 | 1.36 | 0.459 | |
| (3) Y*= 0.07 × RH + 0.05 × Tmin − 1.79 × DPV | 0.61 | −36.72 | 0.00 | 0.492 | |
| Total | (1) Y = 4.087 − 0.037 × RH | 0.24 | −150.00 | 8.13 | 0.079 |
| (2) Y* = 5.195 − 0.051 × RH − 0.995 × DPV | 0.34 | −158.13 | 0.00 | 0.256 | |
| (3) Y*= 4.087 − 0.037 × RH − 0.01 × Tmin − 0.67 × DPV | 0.35 | −158.09 | 0.04 | 0.666 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morffi-Mestre, H.; Ángeles-Pérez, G.; Powers, J.S.; Andrade, J.L.; Huechacona Ruiz, A.H.; May-Pat, F.; Chi-May, F.; Dupuy, J.M. Multiple Factors Influence Seasonal and Interannual Litterfall Production in a Tropical Dry Forest in Mexico. Forests 2020, 11, 1241. https://doi.org/10.3390/f11121241
Morffi-Mestre H, Ángeles-Pérez G, Powers JS, Andrade JL, Huechacona Ruiz AH, May-Pat F, Chi-May F, Dupuy JM. Multiple Factors Influence Seasonal and Interannual Litterfall Production in a Tropical Dry Forest in Mexico. Forests. 2020; 11(12):1241. https://doi.org/10.3390/f11121241
Chicago/Turabian StyleMorffi-Mestre, Hernán, Gregorio Ángeles-Pérez, Jennifer S. Powers, José Luis Andrade, Astrid Helena Huechacona Ruiz, Filogonio May-Pat, Francisco Chi-May, and Juan Manuel Dupuy. 2020. "Multiple Factors Influence Seasonal and Interannual Litterfall Production in a Tropical Dry Forest in Mexico" Forests 11, no. 12: 1241. https://doi.org/10.3390/f11121241