
Annona muricata: Comprehensive Review on the Ethnomedicinal, Phytochemistry, and Pharmacological Aspects Focusing on Antidiabetic Properties
 , , , and
, , , and
Abstract
:1. Introduction
2. Annona Plant
Annona muricata Linn.
3. Ethnomedicinal and Medicinal Uses of A. muricata
3.1. Ethnomedicinal Uses
3.2. Medicinal Uses
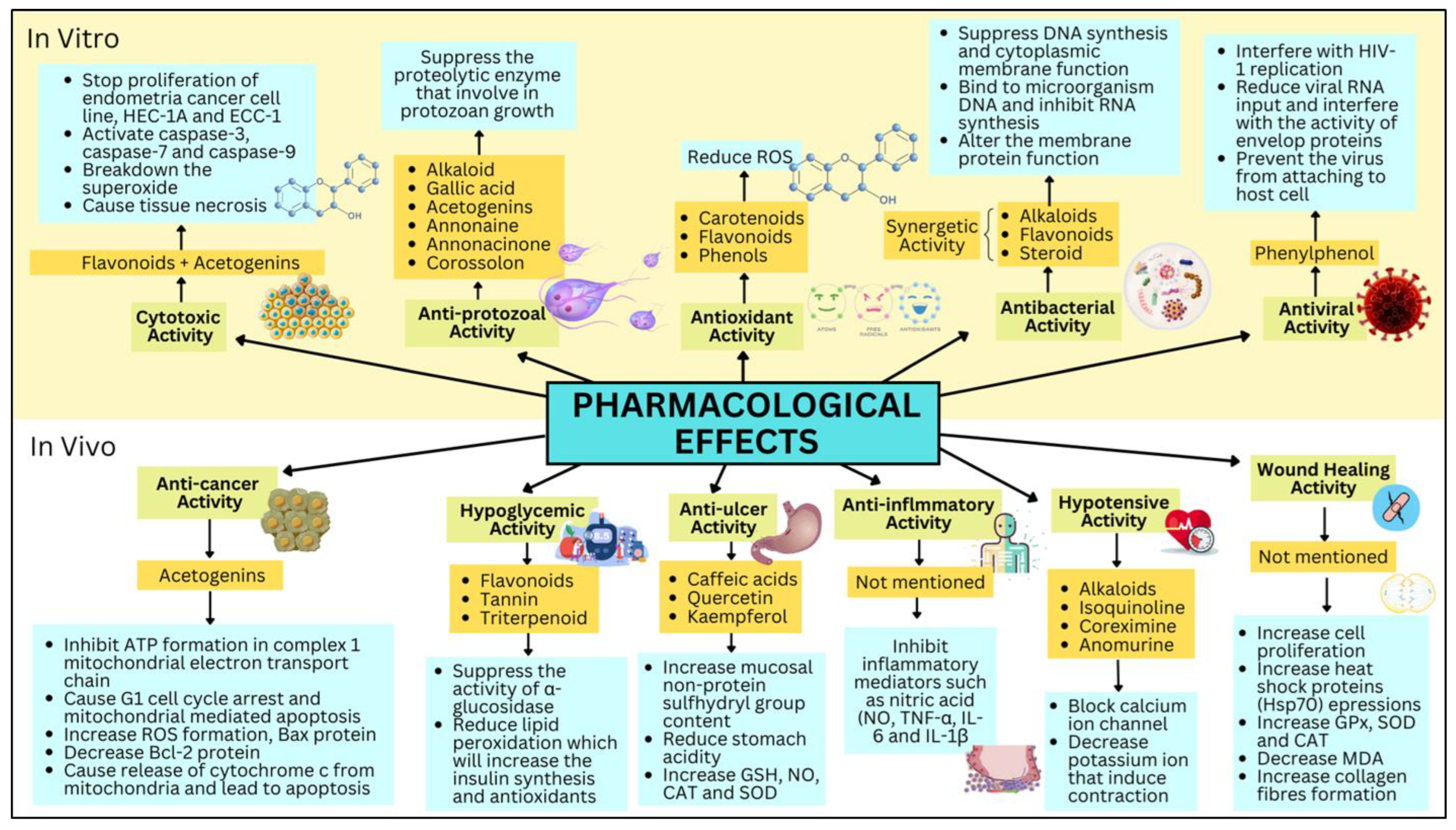
4. Phytochemistry and the Pharmacological Effect of A. muricata
4.1. In Vitro Studies
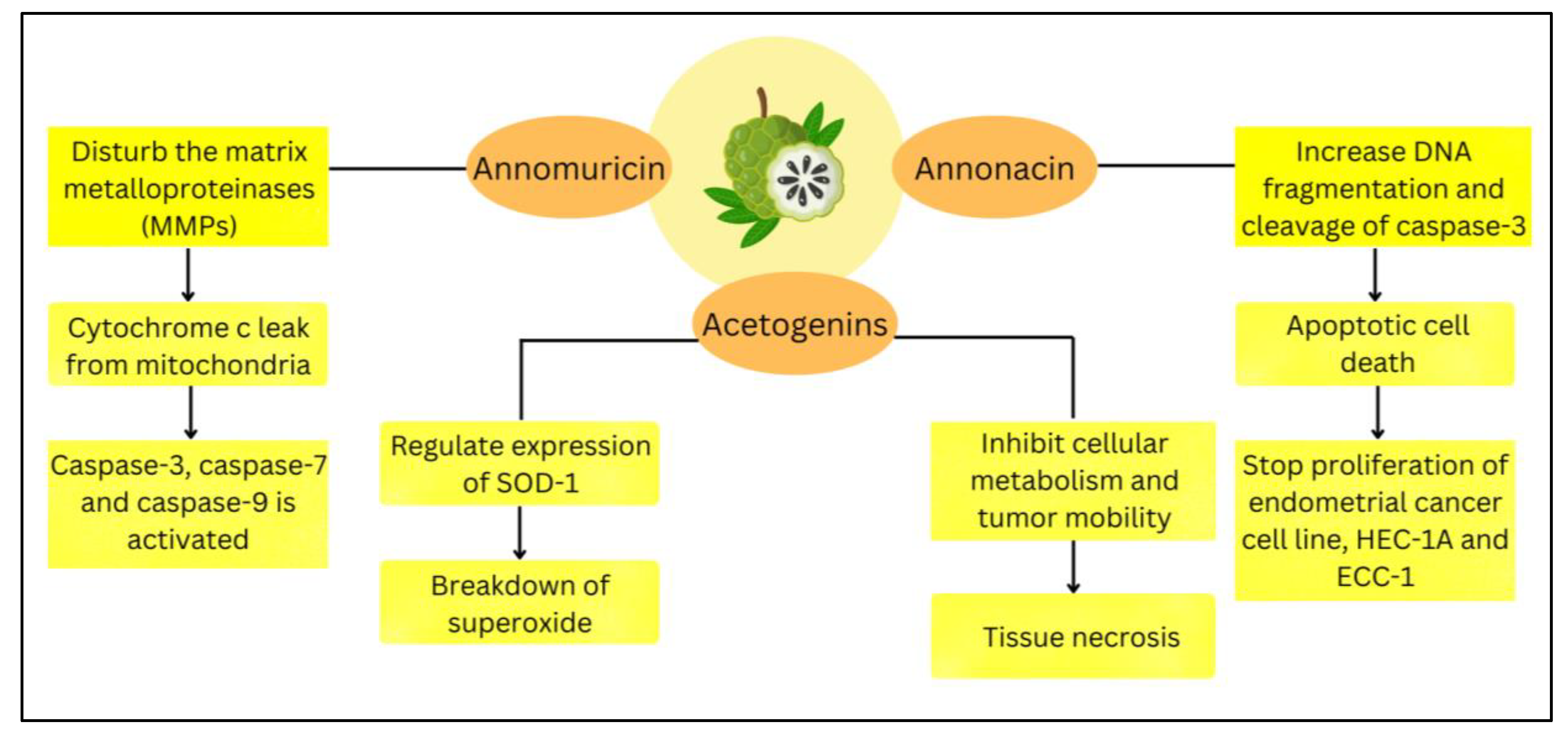
4.1.1. Cytotoxic Activity
4.1.2. Anti-Protozoal Activity
4.1.3. Antioxidant Activity
4.1.4. Anti-Viral Activity
4.2. In Vivo Study
4.2.1. Anti-Cancer Activity
4.2.2. Anti-Ulcer
4.2.3. Anti-Inflammatory Activity
4.2.4. Hypotensive Activity
4.2.5. Wound Healing
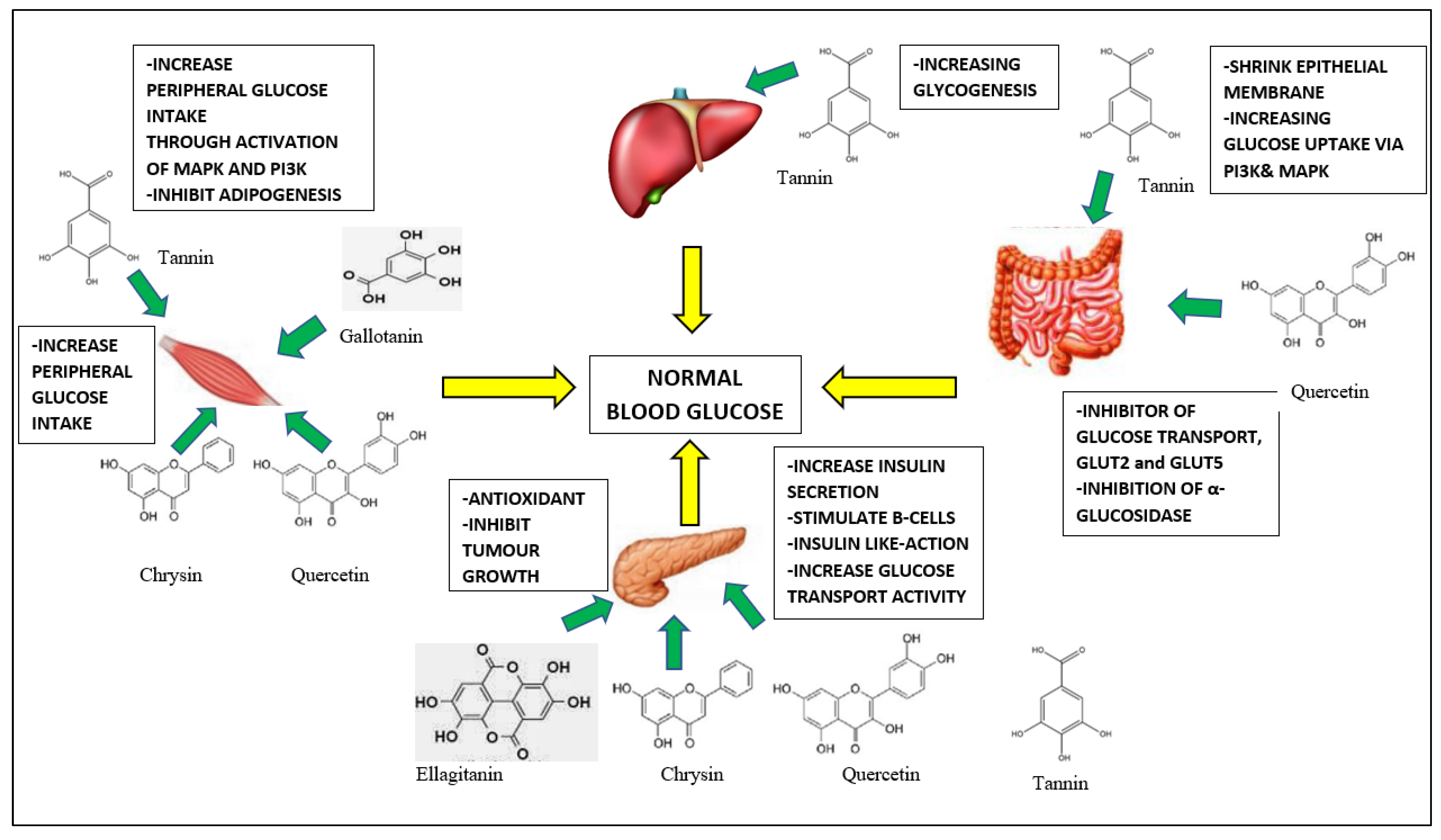
5. Anti-Diabetic Effect of A. muricata
5.1. Hypoglycemic Activity of A. muricata
5.2. In Vivo and In Vitro Study on Anti-Diabetic Effect
6. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ekor, M. The Growing Use of Herbal Medicines: Issues Relating to Adverse Reactions and Challenges in Monitoring Safety. Front. Neurol. 2014, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Moghadamtousi, S.Z.; Fadaeinasab, M.; Nikzad, S.; Mohan, G.; Ali, H.M.; Kadir, H.A. A. muricata (Annonaceae): A Review of Its Traditional Uses, Isolated Acetogenins and Biological Activities. Int. J. Mol. Sci. 2015, 16, 15625–15658. [Google Scholar] [CrossRef]
- Syed Najmuddin, S.U.F.; Romli, M.F.; Hamid, M.; Alitheen, N.B.; Abd Rahman, N.M.A.N. Anti-Cancer Effect of A. muricata Linn Leaves Crude Extract (AMCE) on Breast Cancer Cell Line. BMC Complement. Altern. Med. 2016, 16, 311. [Google Scholar] [CrossRef] [Green Version]
- Miranda, N.C.; Araujo, E.C.B.; Justino, A.B.; Cariaco, Y.; Mota, C.M.; Costa-Nascimento, L.A.; Espindola, F.S.; Silva, N.M. Anti-Parasitic Activity of A. muricata L. Leaf Ethanolic Extract and Its Fractions against Toxoplasma Gondii in Vitro and in Vivo. J. Ethnopharmacol. 2021, 273, 114019. [Google Scholar] [CrossRef]
- Adefegha, S.A.; Oyeleye, S.I.; Oboh, G. Distribution of Phenolic Contents, Antidiabetic Potentials, Antihypertensive Properties, and Antioxidative Effects of Soursop (A. muricata L.) Fruit Parts In Vitro. Biochem. Res. Int. 2015, 2015, 347673. [Google Scholar] [CrossRef] [Green Version]
- Ngemenya, M.N.; Asongana, R.; Zofou, D.; Ndip, R.A.; Itoe, L.O.; Babiaka, S.B. In Vitro Antibacterial Potential against Multidrug-Resistant Salmonella, Cytotoxicity, and Acute Biochemical Effects in Mice of A. muricata Leaf Extracts. Evid. Based Complement. Altern. Med. 2022, 2022, 3144684. [Google Scholar] [CrossRef]
- Paola Balderrama-Carmona, A.; Silva-Beltrán, N.P.; Gálvez-Ruiz, J.-C.; Ruíz-Cruz, S.; Chaidez-Quiroz, C.; Felipe Morán-Palacio, E. Plants Antiviral, Antioxidant, and Antihemolytic Effect of A. muricata L. Leaves Extracts. Plants 2020, 9, 1650. [Google Scholar] [CrossRef]
- Coria-Téllez, A.V.; Montalvo-Gónzalez, E.; Yahia, E.M.; Obledo-Vázquez, E.N. A. muricata: A Comprehensive Review on Its Traditional Medicinal Uses, Phytochemicals, Pharmacological Activities, Mechanisms of Action and Toxicity. Arab. J. Chem. 2018, 11, 662–691. [Google Scholar] [CrossRef] [Green Version]
- Chukwunonso Agu, K.; Eluehike, N.; Oseikhumen Ofeimun, R.; Abile, D.; Ideho, G.; Olukemi Ogedengbe, M.; Omozokpea Onose, P.; Olalekan Elekofehinti, O. Possible Anti-Diabetic Potentials of A. muricata (Soursop): Inhibition of α-Amylase and α-Glucosidase Activities. Clin. Phytoscience 2019, 5, 21. [Google Scholar] [CrossRef]
- Sokpe, A.; Mensah, M.L.K.; Koffuor, G.A.; Thomford, K.P.; Arthur, R.; Jibira, Y.; Baah, M.K.; Adedi, B.; Agbemenyah, H.Y. Hypotensive and Antihypertensive Properties and Safety for Use of A. muricata and Persea Americana and Their Combination Products. Evid. Based Complement. Altern. Med. 2020, 2020, 8833828. [Google Scholar] [CrossRef]
- Opara, P.O.; Enemor, V.H.A.; Eneh, F.U.; Emengaha, F.C. Blood Glucose—Lowering Potentials of A. muricata Leaf Extract in Alloxan—Induced Diabetic Rats. Eur. J. Biol. Biotechnol. 2021, 2, 106–113. [Google Scholar] [CrossRef]
- Banday, M.Z.; Sameer, A.S.; Nissar, S. Pathophysiology of Diabetes: An Overview. Avicenna J. Med. 2020, 10, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and Regional Diabetes Prevalence Estimates for 2019 and Projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th Edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasri, H.; Rafieian-Kopaei, M. Metformin: Current Knowledge. J. Res. Med. Sci. 2014, 19, 658–664. [Google Scholar] [PubMed]
- Florence, N.T.; Benoit, M.Z.; Jonas, K.; Alexandra, T.; Désiré, D.D.P.; Pierre, K.; Théophile, D. Antidiabetic and Antioxidant Effects of A. muricata (Annonaceae), Aqueous Extract on Streptozotocin-Induced Diabetic Rats. J. Ethnopharmacol. 2014, 151, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.S.; Tareq, A.M.; Tareq, S.M.; Farhad, S.; Sayeed, M.A. Screening of Antidiabetic and Antioxidant Potential along with Phytochemicals of Annona Genus: A Review. Futur. J. Pharm. Sci. 2021, 7, 144. [Google Scholar] [CrossRef]
- Mkdt, D.; Abhayagunasekara, A. Selection of Superior Quality Annona Species by Means of Bioactive Compounds and Antioxidant Capacity. World J. Agric. Res. 2020, 8, 39–44. [Google Scholar] [CrossRef]
- Quílez, A.M.; Fernández-Arche, M.A.; García-Giménez, M.D.; De la Puerta, R. Potential Therapeutic Applications of the Genus Annona: Local and Traditional Uses and Pharmacology. J. Ethnopharmacol. 2018, 225, 244–270. [Google Scholar] [CrossRef]
- Leonti, M.; Sahpaz, S.; Karioti, A.; Jantan, I.; Wahab, A.S.; Mariam Abdul Wahab, S.; Areeful Haque, M.; Arshad, L. Exploring the Leaves of A. muricata L. as a Source of Potential Anti-Inflammatory and Anticancer Agents. Front. Pharmacol. 2018, 1, 661. [Google Scholar] [CrossRef]
- Padmanabhan, P.; Paliyath, G. Annonaceous Fruits, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; ISBN 9780123849533. [Google Scholar]
- Kasole, R.; Martin, H.D.; Kimiywe, J. Traditional Medicine and Its Role in the Management of Diabetes Mellitus: “Patients” and Herbalists’ Perspectives. Evid. Based Complement. Altern. Med. 2019, 2019, 2835691. [Google Scholar] [CrossRef]
- Mutakin, M.; Fauziati, R.; Fadhilah, F.N.; Zuhrotun, A.; Amalia, R.; Hadisaputri, Y.E. Pharmacological Activities of Soursop (A. muricata Lin.). Molecules 2022, 27, 1201. [Google Scholar] [CrossRef] [PubMed]
- Adewole, S.O.; Ojewole, J.A.O. Protective Effects of A. muricata Linn. (Annonaceae) Leaf Aqueous Extract on Serum Lipid Profiles and Oxidative Stress in Hepatocytes of Streptozotocin-Treated Diabetic Rats. Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Gavamukulya, Y.; Wamunyokoli, F.; El-Shemy, H.A. A. muricata: Is the Natural Therapy to Most Disease Conditions Including Cancer Growing in Our Backyard? A Systematic Review of Its Research History and Future Prospects. Asian Pac. J. Trop. Med. 2017, 10, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Solanki, J.; Mandaliya, V.B.; Georrge, J.J. Medicinal Properties of A. muricata Extracts in Various Disease. Biochemistry 2020, 2020, 126–133. [Google Scholar]
- Satia, A.N.; Damayanti, Y.D.; Wangchuk, P.; Keller, P.A. Alkaloid Diversity, and Pharmacological Activities. Molecules 2019, 24, 4419. [Google Scholar]
- Vijayameena, C.; Subhashini, G.; Loganayagi, M.; Ramesh, B. Phytochemical Screening and Assessment of Antibacterial Activity for the Bioactive Compounds in A. muricata. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 1–8. [Google Scholar]
- Kim, G.T.; Tran, N.K.S.; Choi, E.H.; Song, Y.J.; Song, J.H.; Shim, S.M.; Park, T.S. Immunomodulatory Efficacy of Standardized A. muricata (Graviola) Leaf Extract via Activation of Mitogen-Activated Protein Kinase Pathways in RAW 264.7 Macrophages. Evid. Based Complement. Altern. Med. 2016, 2016, 2905127. [Google Scholar] [CrossRef] [Green Version]
- Afzaal, M.; Saeed, F.; Asghar, A.; Shah, Y.A.; Ikram, A.; Ateeq, H.; Hussain, M.; Ofoedu, C.E.; Chacha, J.S. Review Article Nutritional and Therapeutic Potential of Soursop. J. Food Qual. 2022, 2022, 8828358. [Google Scholar] [CrossRef]
- Riley-Saldaña, C.A.; Cruz-Ortega, M.D.R.; Martínez Vázquez, M.; De-La-Cruz-Chacón, I.; Castro-Moreno, M.; González-Esquinca, A.R. Acetogenins and Alkaloids during the Initial Development of A. muricata L. (Annonaceae). Z. Fur Naturforsch. Sect. C J. Biosci. 2017, 72, 497–506. [Google Scholar] [CrossRef]
- Ranisaharivony, B.G.; Ramanandraibe, V.; Rasoanaivo, L.H.; Rakotovao, M.; Lemaire, M. Separation and Potential Valorization of Chemical Constituents of Soursop Seeds. J. Pharmacogn. Phytochem. 2015, 4, 161–171. [Google Scholar]
- Toto Blessing, L.D.; Colom, O.Á.; Popich, S.; Neske, A.; Bardón, A. Antifeedant and Toxic Effects of Acetogenins from A. Montana on Spodoptera Frugiperda. J. Pest Sci. 2010, 83, 307–310. [Google Scholar] [CrossRef]
- Wu, F.E.; Gu, Z.M.; Zeng, L.; Zhao, G.X.; Zhang, Y.; McLaughlin, J.L.; Sastrodihardjo, S. Two New Cytotoxic Monotetrahydrofuran Annonaceous Acetogenins, Annomuricins A and B, from the Leaves of A. Muricata. J. Nat. Prod. 1995, 58, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Liu, J.; Sun, X.; Zhu, W.; Yang, F.; Felczak, L.; Ping Dou, Q.; Zhou, K. Novel Annonaceous Acetogenins from Graviola (A. Muricata) Fruits with Strong Anti-Proliferative Activity. Tetrahedron Lett. 2017, 58, 1895–1899. [Google Scholar] [CrossRef]
- Kim, G.S.; Zeng, L.; Alali, F.; Rogers, L.L.; Wu, F.E.; McLaughlin, J.L.; Sastrodihardjo, S. Two New Mono-Tetrahydrofuran Ring Acetogenins, Annomuricin E and Muricapentocin, from the Leaves of A. Muricata. J. Nat. Prod. 1998, 61, 432–436. [Google Scholar] [CrossRef]
- Moghadamtousi, S.Z.; Rouhollahi, E.; Karimian, H.; Fadaeinasab, M.; Firoozinia, M.; Abdulla, M.A.; Kadir, H.A. The Chemopotential Effect of A. muricata Leaves against Azoxymethane-Induced Colonic Aberrant Crypt Foci in Rats and the Apoptotic Effect of Acetogenin Annomuricin E in HT-29 Cells: A Bioassay-Guided Approach. PLoS ONE 2015, 10, e0122288. [Google Scholar] [CrossRef]
- Zeng, L.; Wu, F.-E.; Oberlies, N.H.; Mclaughlin, J.L.; Sastrodihadjo, S. Five New Monotetrahydrofuran Ring Acetogenins from the Leaves of A. Muricata. J. Nat. Prod. 1996, 59, 1035–1042. [Google Scholar] [CrossRef]
- Wu, F.-E.; Zhao, G.-X.; Zheng, L.; Zhang, Y.; Schwedler, J.T.; McLaughlin, J.L. Additional Bioactive Acetogenins, Annomutacin and (2,4-Trans and Cis)-10r-Annonacin-A-Ones, from The Leaves of A. Muricata. J. Nat. Prod. 1995, 58, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Wu, F.-E.; McLaughlin, J. Annohexocin, A Novel Mono-THF Acetogenin with Six Hydroxyls, from A. muricata (Annonaceae). Bioorg. Med. Chem. Lett. 1995, 5, 1865–1868. [Google Scholar] [CrossRef]
- Rieser, M.J.; Gu, Z.; Fang, X.; Zeng, L.; Wood, K.V.; Mclaughlin, J.L. Five Novel Mono-Tetrahydrofuran Ring Acetogenins from the Seeds of A. Muricata. J. Nat. 1996, 59, 100–108. [Google Scholar] [CrossRef]
- Prasad, S.K.; Pradeep, S.; Shimavallu, C.; Kollur, S.P.; Syed, A.; Marraiki, N.; Egbuna, C.; Gaman, M.-A.; Kosakowska, O.; Cho, W.C.; et al. Evaluation of A. Muricata Acetogenins as Potential Anti-SARS-CoV-2 Agents Through Computational Approaches. Front. Chem. 2021, 8, 624716. [Google Scholar] [CrossRef]
- Jeevitha Priya, M.; Vidyalakshmi, S.; Rajeswari, M. Study on Reversal of ABCB1 Mediated Multidrug Resistance in Colon Cancer by Acetogenins: An in-Silico Approach. J. Biomol. Struct. Dyn. 2022, 40, 4273–4284. [Google Scholar] [CrossRef] [PubMed]
- Liaw, C.C.; Chang, F.R.; Lin, C.Y.; Chou, C.J.; Chiu, H.F.; Wu, M.J.; Wu, Y.C. New Cytotoxic Monotetrahydrofuran Annonaceous Acetogenins from A. Muricata. J. Nat. Prod. 2002, 65, 470–475. [Google Scholar] [CrossRef]
- Chang, F.R.; Liaw, C.C.; Lin, C.Y.; Chou, C.J.; Chiu, H.F.; Wu, Y.C. New Adjacent Bis-Tetrahydrofuran Annonaceous Acetogenins from A. Muricata. Planta Med. 2003, 69, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Vila-Nova, N.S.; de Morais, S.M.; Falcão, M.J.C.; Alcantara, T.T.N.; Ferreira, P.A.T.; Cavalcanti, E.S.B.; Vieira, I.G.P.; Campello, C.C.; Wilson, M. Different Susceptibilities of Leishmania Spp. Promastigotes to the A. Muricata Acetogenins Annonacinone and Corossolone, and the Platymiscium Floribundum Coumarin Scoparone. Exp. Parasitol. 2013, 133, 334–338. [Google Scholar] [CrossRef] [PubMed]
- De Pedro, N.; Cautain, B.; Melguizo, A.; Cortes, D.; Vicente, F.; Genilloud, O.; Tormo, J.R.; Peláez, F. Analysis of Cytotoxic Activity at Short Incubation Times Reveals Profound Differences among Annonaceus Acetogenins, Inhibitors of Mitochondrial Complex I. J. Bioenerg. Biomembr. 2013, 45, 145–152. [Google Scholar] [CrossRef]
- Pautus, S.; Alami, M.; Adam, F.; Bernadat, G.; Lawrence, D.A.; De Carvalho, A.; Ferry, G.; Rupin, A.; Hamze, A.; Champy, P.; et al. Characterization of the Annonaceous Acetogenin, Annonacinone, a Natural Product Inhibitor of Plasminogen Activator Inhibitor-1. Sci. Rep. 2016, 6, 36462. [Google Scholar] [CrossRef] [Green Version]
- Hopp, D.C.; Zeng, L.; Gu, Z.M.; Kozlowski, J.F.; McLaughlin, J.L. Novel Mono-Tetrahydrofuran Ring Acetogenins, from the Bark of A. Squamosa, Showing Cytotoxic Selectivities for the Human Pancreatic Carcinoma Cell Line, PACA-2. J. Nat. Prod. 1997, 60, 581–586. [Google Scholar] [CrossRef]
- Landolt, J.L.; Ahammadsahib, K.I.; Hollingworth, R.M.; Barr, R.; Crane, F.L.; Buerckv, N.L.; McCabe, G.P.; McLaughlin, J.L. Determination of Structure-Activity Relationships of Annonaceous acetogenins by Inhibition of Oxygen Uptake in Rat Liver Mitochondria. Chem. Biol. Interact. 1995, 98, 1–13. [Google Scholar] [CrossRef]
- Liaw, C.C.; Chang, F.R.; Wu, C.C.; Chen, S.L.; Bastow, K.F.; Hayashi, K.I.; Nozaki, H.; Lee, K.H.; Wu, Y.C. Nine New Cytotoxic Monotetrahydrofuranic Annonaceous Acetogenins from Annona montana. Planta Med. 2004, 70, 948–959. [Google Scholar] [CrossRef]
- Ravi, L.; Bhardwaj, V.; Venkatraman, M.; Gopiesh Khanna, V. Annoreticuin and Sabadelin, a Potential Oncogenic Transcriptional Factor Inhibitors: An in Silico Analysis. Der Pharm. Lett. 2015, 7, 204–212. [Google Scholar]
- Gleye, C.; Laurens, A.; Hocquemiller, R.; Laprevote, O.; Serani, L. Cohibins A and B, Acetogenins from Roots of A. muricata. Phytochemistry 1997, 44, 1541–1545. [Google Scholar] [CrossRef]
- Gleye, C.; Raynaud, S.; Fourneau, C.; Laurens, A.; Laprévote, O.; Serani, L.; Fournet, A.; Hocquemiller, R. Cohibins C and D, Two Important Metabolites in the Biogenesis of Acetogenins from A. Muricata and Annona Nutans. J. Nat. Prod. 2000, 63, 1192–1196. [Google Scholar] [CrossRef] [PubMed]
- Cavé, A.; Figadère, B.; Laurens, A.; Cortes, D. Acetogenins from Annonaceae. Fortschr Chem Org Naturst. 1997, 70, 81–288. [Google Scholar] [CrossRef] [PubMed]
- Gromek, D.; Figadère, B.; Hocquemiller, R.; Cavé, A.; Cortes, D. Corepoxylone, a Possible Precursor of Mono-Tetrahydrofuran γ-Lactone Acetogenins: Biomimetic Synthesis of Corossolone. Tetrahedron 1993, 49, 5247–5252. [Google Scholar] [CrossRef]
- Yu, D.Q. Recent Works on Anti-Tumor Constituent from Annonaceae Plants in China. Pure Appl. Chem. 1999, 71, 1119–1122. [Google Scholar] [CrossRef]
- Zhao, G.X.; Rieser, M.J.; Hui, Y.H.; Miesbauer, L.R.; Smith, D.L.; McLaughlin, J.L. Biologically Active Acetogenins from Stem Bark of Asimina Triloba. Phytochemistry 1993, 33, 1065–1073. [Google Scholar] [CrossRef]
- Xu, L.; Chang, C.J.; Yu, J.G.; Cassady, J.M. Chemistry and Selective Cytotoxicity of Annonacin-10-One, Isoannonacin, and Isoannonacin-10-One: Novel Polyketides from Annona Densicoma (Annonaceae). J. Org. Chem. 1989, 54, 5418–5421. [Google Scholar] [CrossRef]
- Chang, F.R.; Wu, Y.C. Novel Cytotoxic Annonaceous Acetogenins from A. Muricata. J. Nat. Prod. 2001, 64, 925–931. [Google Scholar] [CrossRef]
- Wu, F.-E.; Zheng, L.; Gu, Z.-M.; Zhao, G.-X.; Zhang, Y.; Schwendler, J.T.; McLaughlin, J.L. Muricatocins A And B, Two New Bioactive Monotetrahydrofuran Annonaceous Acetogenins From The Leaves of A. Muricata. J. Nat. Prod. 1995, 58, 902–908. [Google Scholar] [CrossRef]
- Wu, F.E.; Zeng, L.; Gu, Z.M.; Zhao, G.X.; Zhang, Y.; Schwedler, J.T.; McLaughlin, J.L.; Sastrodihardjo, S. New Bioactive Monotetrahydrofuran Annonaceous Acetogenins, Annomuricin C and Muricatocin C, from the Leaves of A. Muricata. J. Nat. Prod. 1995, 58, 909–915. [Google Scholar] [CrossRef]
- Sun, S.; Liu, J.; Kadouh, H.; Sun, X.; Zhou, K. Three New Anti-Proliferative Annonaceous Acetogenins with Mono-Tetrahydrofuran Ring from Graviola Fruit (A. Muricata). Bioorg. Med. Chem. Lett. 2014, 24, 2773–2776. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.S.; Zeng, L.; Alali, F.; Rogers, L.L.; Wu, F.E.; Sastrodihardjo, S.; McLaughlin, J.L. Muricoreacin and Murihexocin C, Mono Tetrahydrofuran Acetogenins, from the Leaves of A. Muricata. Phytochemistry 1998, 49, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Dzhemilev, U.M.; D’Yakonov, V.A.; Tuktarova, R.A.; Dzhemileva, L.U.; Ishmukhametova, S.R.; Yunusbaeva, M.M.; De Meijere, A. Short Route to the Total Synthesis of Natural Muricadienin and Investigation of Its Cytotoxic Properties. J. Nat. Prod. 2016, 79, 2039–2044. [Google Scholar] [CrossRef]
- Gleye, C.; Raynaud, S.; Hocquemiller, R.; Laurens, A.; Fourneau, C.; Serani, L.; Laprévote, O.; Roblot, F.; Leboeuf, M.; Fournet, A.; et al. Muricadienin, Muridienins and Chatenaytrienins, the Early Precursors of Annonaceous Acetogenins. Phytochemistry 1998, 47, 749–754. [Google Scholar] [CrossRef]
- Rieser, M.J.; Kozlowski, J.F.; Wood, K.V.; McLaughlin, J.L. Muricatacin: A Simple Biologically Active Acetogenin Derivative from the Seeds of A. Muricata (Annonaceae). Tetrahedron Lett. 1991, 32, 1137–1140. [Google Scholar] [CrossRef]
- Sun, S.; Liu, J.; Zhou, N.; Zhu, W.; Dou, Q.P.; Zhou, K. Isolation of Three New Annonaceous Acetogenins from Graviola Fruit (A. Muricata) and Their Anti-Proliferation on Human Prostate Cancer Cell PC-3. Bioorg. Med. Chem. Lett. 2016, 26, 4382–4385. [Google Scholar] [CrossRef]
- Yu, J.G.; Gui, H.Q.; Luo, X.Z.; Sun, L. Murihexol, a Linear Acetogenin from A. Muricata. Phytochemistry 1998, 49, 1689–1692. [Google Scholar] [CrossRef]
- Calzada, F.; Merlin-Lucas, V.I.; Valdes, M.; Solares-Pascasio, J.I.; Garcia-Hernandez, N.; Pina-Jimenez, E.; Velazquez, C.; Barbosa, E.; Yepez-Mulia, L.; Ordoñez-Razo, R.M. Secondary Metabolites and Biological Properties of A. muricata. Rev. Bras. Farmacogn. 2020, 30, 305–311. [Google Scholar] [CrossRef]
- Fofana, S.; Ziyaev, R.; Abdusamatov, A.; Zakirov, S.K. Alkaloids from A. Muricata Leaves. Chem. Nat. Compd. 2011, 47, 321. [Google Scholar] [CrossRef]
- Hasrat, J.A.; De Bruyne, T.; De Backer, J.P.; Vauquelin, G.; Vlietinck, A.J. Isoquinoline Derivatives Isolated from the Fruit of A. Muricata as 5-HTergic 5-HT(1A) Receptor Agonists in Rats: Unexploited Antidepressive (Lad) Products. J. Pharm. Pharmacol. 1997, 49, 1145–1149. [Google Scholar] [CrossRef]
- Matsushige, A.; Kotake, Y.; Matsunami, K.; Otsuka, H.; Ohta, S.; Takeda, Y. Annonamine, a New Aporphine Alkaloid from the Leaves of A. Muricata. Chem. Pharm. Bull. 2012, 60, 257–259. [Google Scholar] [CrossRef] [Green Version]
- Leboeuf, M.; Legueut, C.; Cavé, A.; Desconclois, J.F.; Forgacs, P.; Jacquemin, H. Alcaloides Des Annonacées XXIX1: Alcaloides de l’A. Muricata L. J. Med. Plant Res. 1981, 42, 37–44. [Google Scholar] [CrossRef]
- Fofana, S.; Keita, A.; Balde, S.; Ziyaev, R.; Aripova, S.F. Alkaloids from Leaves of A. Muricata. Chem. Nat. Compd. 2012, 48, 714. [Google Scholar] [CrossRef] [Green Version]
- Mohantya, S.; Hollinshead, J.; Jones, L.; Jones, P.W.; Thomas, D.; Alison, A.; Watson, D.G.; Gray, A.I.; Molyneux, R.J.; Nash, R.J. A. Muricata (Graviola): Toxic or Therapeutic Sambeet. Nat. Prod. Commun. 2010, 1, 9–12. [Google Scholar]
- Cijo George, V.; Naveen Kumar, D.R.; Rajkumar, V.; Suresh, P.K.; Ashok Kumar, R. Quantitative Assessment of the Relative Antineoplastic Potential of the N-Butanolic Leaf Extract of A. Muricata Linn. in Normal and Immortalized Human Cell Lines. Asian Pac. J. Cancer Prev. 2012, 13, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Nawwar, M.; Ayoub, N.; Hussein, S.; Hashim, A.; El-Sharawy, R.; Wende, K.; Harms, M.; Lindequist, U. A Flavonol Triglycoside and Investigation of the Antioxidant and Cell Stimulating Activities of A. Muricata Linn. Arch. Pharm. Res. 2012, 35, 761–767. [Google Scholar] [CrossRef]
- Marques, V.; Farah, A. Chlorogenic Acids and Related Compounds in Medicinal Plants and Infusions. Food Chem. 2009, 113, 1370–1376. [Google Scholar] [CrossRef]
- George, V.C.; Kumar, D.R.N.; Suresh, P.K.; Kumar, R.A. Antioxidant, DNA Protective Efficacy and HPLC Analysis of A. Muricata (Soursop) Extracts. J. Food Sci. Technol. 2015, 52, 2328–2335. [Google Scholar] [CrossRef]
- Leesombun, A.; Boonmasawai, S.; Nishikawa, Y. Ethanol Extracts from Thai Plants Have Anti-Plasmodium and Anti-Toxoplasma Activities In Vitro. Acta Parasitol. 2019, 64, 257–261. [Google Scholar] [CrossRef]
- Rady, I.; Bloch, M.B.; Chamcheu, R.C.N.; Banang Mbeumi, S.; Anwar, M.R.; Mohamed, H.; Babatunde, A.S.; Kuiate, J.R.; Noubissi, F.K.; El Sayed, K.A.; et al. Anticancer Properties of Graviola (A. Muricata): A Comprehensive Mechanistic Review. Oxid. Med. Cell. Longev. 2018, 2018, 1826170. [Google Scholar] [CrossRef] [Green Version]
- Carmona, A.P.B.; Beltran, N.P.S.; Ruiz, J.-C.G.; Ruiz-Cruz, S.; Quiroz, C.C.; Palacio, E.F.M. Antiviral, Antioxidant, and Antihemolytic Effect of A. muricata L. Leaves Extracts. Plants 2020, 9, 1650. [Google Scholar] [CrossRef] [PubMed]
- Indrawati, L.; Bela, B.; Abdullah, M.; Surono, I.S.; Indrawati, L. The Effect of an A. Muricata Leaf Extract on Nutritional Status and Cytotoxicity in Colorectal Cancer: A Randomized Controlled Trial. Asia Pac. J. Clin. Nutr. 2016, 26, 606–612. [Google Scholar] [CrossRef]
- Nwokocha, C.R.; Owu, D.U.; Gordon, A.; Thaxter, K.; Mccalla, G.; Ozolua, R.I.; Young, L. Possible Mechanisms of Action of the Hypotensive Effect of A. Muricata (Soursop) in Normotensive Sprague-Dawley Rats. Pharm. Biol. 2012, 50, 1436–1441. [Google Scholar] [CrossRef] [Green Version]
- Moghadamtousi, S.Z.; Rouhollahi, E.; Hajrezaie, M.; Karimian, H.; Abdulla, M.A.; Kadir, H.A. A. Muricata Leaves Accelerate Wound Healing in Rats via Involvement of Hsp70 and Antioxidant Defence. Int. J. Surg. 2015, 18, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, D.O.; Komolafe, O.A.; Adewole, O.S.; Martins, E.M.; Kehinde, A.T. Anti Hyperglycemic Activities of A. Muricata (Linn). Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 62. [Google Scholar] [CrossRef] [Green Version]
- Graf, B.A.; Milbury, P.E.; Blumberg, J.B. Flavonols, Flavones, Flavanones, and Human Health: Epidemiological Evidence. J. Med. Food 2005, 8, 281–290. [Google Scholar] [CrossRef]
- Vinayagam, R.; Xu, B. Antidiabetic Properties of Dietary Flavonoids: A Cellular Mechanism Review. Nutr. Metab. 2015, 12, 60. [Google Scholar] [CrossRef] [Green Version]
- Son, Y.; Lee, H.; Son, S.Y.; Lee, C.H.; Kim, S.Y.; Lim, Y. Ameliorative Effect of A. Muricata (Graviola) Extract on Hyperglycemia Induced Hepatic Damage in Type 2 Diabetic Mice. Antioxidants 2021, 10, 1546. [Google Scholar] [CrossRef]
- Setiadi, R.R.; Zein, A.F.M.Z.; Nauphar, D. Antihyperglicemic Effectiveness Comparison of Ethanol Extract of Soursop Leaf (A. Muricata L.) Againts Acarbose in Streptozotocin-Induced Diabetic White Rats. J. Phys. Conf. Ser. 2019, 1146, 012009. [Google Scholar] [CrossRef]
- Sovia, E.; Ratwita, W.; Wijayanti, D.; Novianty, D.R.; Yani, A.; Terusan Jenderal Sudirman, J.; Java, W. Hypoglycemic and Hypolipidemic Effects of A. Muricata L. Leaf Ethanol Extract. Int. J. Pharm. Pharm. Sci. 2017, 9, 170–174. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Compound | Part of Plant | Activity | Effect |
|---|---|---|---|
| Acetogenins | |||
| 1. Annomuricin (annomuricin A, B, C) [2,33,34] | Leaves, pericarp | Cytotoxic [33] | Annomuricin A showed toxicity against human breast carcinoma (MCF-7) showed ED50 value of >1.0 μg/mL, human colon adenocarcinoma (HT-29) with ED50 >1.0 μg/mL, human lung cancer (A549) with ED50 of 3.30 × 10−1 μg/mL and brine shrimp (BST) LC50 result showed value of 6.25 × 10−1 μg/mL [33]. On the other hand, annomuricin B revealed toxicity against human breast carcinoma (MCF-7) with ED50 value of >1.0 μg/mL, human colon adenocarcinoma (HT-29) with ED50 of 4.35 × 10−1 μg/mL, human lung cancer (A549) with ED50 of 1.59 × 10−1 μg/mL and brine shrimp (BST) LC50 result showed value of 6.87 × 10−1 μg/mL [33]. Moreover, treatment of annonamuricin A, B and C at dose 20 μg/mL reduced PC-3 cell viability by 86.0, 96.9, and 97.7% respectively [34]. |
| 2. Annomuricin E [35,36] | Leaves | Cytotoxic [36] | Toxicity towards colon HT-29 cancer cells with ED50 values of 6.68 × 10−2 μg/mL and pancreatic carcinoma PACA-2 with ED50 of 2.42 × 10−2 μg/mL [35]. Besides, IC50 dose of compounds against HT-29 cells after 12, 24 and 48 hrs of treatment showed a result of 5.72 ± 0.41 μg/mL, 3.49 ± 0.22 μg/mL and 1.62 ± 0.24 μg/mL respectively [36]. |
| 3. Cis-annomuricin-D-one, trans-annomuricin-D-one [37] | Leaves | Cytotoxic [37] | Toxicity against lung cancer cells A549 with ED50 of <10−2 μg/mL, colon HT-29 with ED50 of <10−2 μg/mL, and pancreatic PACA-2 with ED50 of <10−2 μg/mL [37]. |
| 4. Annomutacin [38] | Leaves | Cytotoxic [38] | Toxicity against lung A549, breast MCF-7 and colon HT-29 with ED50 values of 1.57 × 10−2, >1.0 and >1.0 μg/mL respectively [38]. |
| 5. Annohexocin [39] | Leaves | Cytotoxic [39] | It showed selective toxicity in lung A549 with ED50 of 0.34 μg/mL, pancreatic PACA-2 with ED50 of 0.77, colon HT-29 with ED50 of 0.78, and breast MCF-7 with ED50 of 2.26 μg/mL [39]. |
| 6. Annonacin [34] | Seeds, leaves, pericarp | Cytotoxic, insecticidal, anti-microbial, anti-tumor, neurotoxic, neurodegenerative [34] | Anti-proliferative effect on PC-3 cell which the cell viability was decreased by 96.9% with dose at 20 μg/mL [34]. |
| 7. Cis-annonacin [40] | Seeds | Cytotoxic [40] | Crown gall tumour inhibition (28%), brine shrimp toxicity (LC50 of 2.3 μg/mL), lung A549 (LC50 of 2.3 μg/mL), breast MCF-7 (IC50 of 1.18 μg/mL), and colon HT-29 cancer cell toxicity (IC50 of 1.0 × 10−8 μg/mL) [40]. |
| 8. Annonacin-10-one [41] | Seeds | Cytotoxic, anti-viral [41] | Able to prevent breast cancer activity and halt the SARS-CoV-2 spike protein from relocating [41]. |
| 9. Cis-annonacin-10-one [40] | Seeds | Cytotoxic [40] | Crown gall tumour inhibition (32%), brine shrimp toxicity (LC50 of 1.8 μg/mL), lung A549 (IC50 of 3.5 × 10−1 μg/mL), breast MCF-7 (IC50 of 2.9 × 10−1 μg/mL), and colon HT-29 cancer cell toxicity (IC50 of 9.0 × 10−4 μg/mL) [40]. |
| 10. (2,4-cis)-10R-annonacin-A-one, (2,4-trans)-10R-annonacin-A-one [38] | Leaves | Cytotoxic [38] | Toxicity against colon HT-29, breast carcinoma MCF-7 and ling A549 with results of ED50 of >1.0, 5.70 × 10−1, and 1.74 × 10−1 μg/mL respectively [38]. |
| 11. Annonacin A [42] | Leaves, seeds, pericarp | Cytotoxic [42] | Could possibly reverse MDR, which is caused by ABCB1 in colorectal cancer. This would make it possible for the tested anti-cancer drugs to work better against tumours [42]. |
| 12. Annopentocin A, B, C [37] | Leaves | Cytotoxic [37] | Annopentocin A, B and C have toxicity against lung A549 with ED50 of 1.71 × 10−1, 2.74 × 10−2 and 2.06 × 10−2 μg/mL respectively [37]. Besides, these compounds also have effect on colon HT-29 (1.63, 1.64, and 1.24 μg/mL) and pancreatic cancer cells PACA-2 (3.58 × 10−2, 1.62 × 10−1, and 4.28 × 10−3 μg/mL) [37]. |
| 13. Annocatalin [43] | Leaves | Cytotoxic [43] | Toxicity in the presence of two types of human hepatoma cells, hep G2 and hep 2,2,15 which the IC50 result were 5.70 and 3.48 × 10−3 μg/mL respectively [43]. |
| 14. Annocatacin A [44] | Seeds | Cytotoxic [44] | Toxicity in the presence of hepatoma hep G2 and hep 2,2,15 with IC50 of 12.11 and 8.17 × 10−1 μg/mL respectively [44]. |
| 15. Annocatacin B [44] | Leaves | Cytotoxic [44] | Toxicity in the presence of hepatoma hep G2 and hep 2,2,15 with IC50 of 3.35 × 10−2 and 2.22 × 10−1 μg/mL respectively [44]. |
| 16. Arianacin [40] | Seeds | Cytotoxic [40] | Crown gall tumour inhibition (26%), brine shrimp toxicity (LC50 of 7.1 μg/mL), lung A549 (IC50 of 4.7 × 10−3 μg/mL), breast MCF-7 (IC50 of 4.0 × 10−1 µg/m), and colon HT-29 cancer cell toxicity (IC50 of 4.4 μg/mL) [40]. |
| 17. Cis-annomontacin [43] | Seeds | Cytotoxic [43] | Toxicity in the presence of two hepatoma cell, hep G2 and hep 2,2,15 which the IC50 value showed values of 2.98 × 10−1 and 1.62 × 10−2 μg/mL [43]. |
| 18. Annonacinone [45] | Seeds, leaves | Cytotoxic, inhibitory activity, leishmanicidal activity [45,46,47] | Annonacinone enhances the fibrinolytic effect of tissue plasminogen activator (tPA). Annonacinone decreased PAI-1/tPA complex formation by enhancing the substrate pathway [47]. Leishmanicidal activity against Leishmania species (L. donovani, L. major and L. mexicana) which the value of IC50, 7.66 ± 0.77, 6.72 ± 0.37, and 8.00 ± 1.00 μg/mL respectively [45]. Moreover, also capable to cause toxicity in lung A549 (126 ± 44 μg/mL), hepatoma hep G2 (20 ± 10 μg/mL) and colon Ht-29 (74 ± 7 μg/mL) [46]. |
| 19. Annoreticuin-9-one [48] | Seeds | Cytotoxic [48] | Cytotoxic activities against brine shrimp lethality test (BST) with LC50 value of 2.4 × 10–4 μg/mL, ED50 for pancreatic tumour cell line (PACA-2) was 2.4 × 10–4 μg/mL; human lung carcinoma (A-549) with ED50 = 2.7 × 10–1 μg/mL, human prostate cancer (PC-3) showed ED50 value of 9.8 × 10–3 μg/mL and ED50 for human lung carcinoma (A-549) was 2.7 × 10–1 μg/mL [48]. |
| 20. Asimicin [49] | Leaves | Cytotoxic [49] | Toxicity on HT-29 human colon cancer cell line (ED50 of 3.3 × 10−11 μg/mL) and lung A549 (10−3 μg/mL) [49]. |
| 21. Bullatacin [44] | Seeds | Cytotoxic [44] | Toxicity against hepatoma hep G2 and hep 2,2,15 with IC50 values of 6.30 × 10−5 and 6.90 × 10−5 μg/mL respectively [44]. |
| 22. Bullatalicin [49] | Seeds | Cytotoxic [49] | Toxicity against lung A549 (ED50 of 2.34 × 10−7 μg/mL), breast MCF-7 (ED50 of 2.34 μg/mL), colon HT-29 (ED50 8.8 × 10−1 μg/mL), and brine shrimp test BST (LC50 of 45.56 μg/mL) [49]. |
| 23. Cis-annoreticuin [50] | Fruits | Cytotoxic [50] | Toxicity on human hepatoma carcinoma cell line (HepG2) with ED50 value of 2.4 × 10–3 μg/mL [50]. |
| 24. (Sabadelin) chatenaytrienin 1, 2, 3 [51] | Roots | Anti-tumor, cytotoxic [49] | Inhibition of T-Box transcriptional factor (TBX5) and Murine Double Minute 2 (MDM2) [49]. |
| 25. Cohibin A, B [52] | Roots, seeds | NR | NR |
| 26. Cohibin C, D [53] | Seeds | NR | NR |
| 27. Corossolone [45] | Leaves, seeds | Cytotoxic [45] | Anti-leishmanial activity with EC50 value of between 16.14–18.73 μg/mL [45]. In addition, toxicity against oral KB cancer cells (1 × 10−1 μg/mL) [54]. |
| 28. Cis-corossolone [43] | Leaves | Cytotoxic [43] | Toxicity in the presence of two hepatoma cell, hep G2 and hep 2,2,15 which the IC50 value showed values of 1.65 × 10−1 and 4.76 × 10−2 μg/mL [43]. |
| 29. Corossolin [54] | Seeds, leaves | Cytotoxic [54] | Cytotoxic activity against oral KB cancer cells (ED50 of 3 × 10−3 μg/mL) and VERO cells (ED50 of 3 × 10−2 μg/mL) [54]. |
| 30. Corepoxylone [55] | Seeds | NR | NR |
| 31. Donhexocin [56] | Seeds | Anti-tumour, cytotoxic [56] | In vitro inhibition of human leukaemia (HL-60) and human colon adenocarcinoma (HCT-8) cell lines with IC50 of <1 μg/mL [56]. |
| 32. Gigantetronenin [32] | Leaves, seeds | Cytotoxic, insecticides [32] | Inhibit NADH oxidase (IC50 of 3.7 ± 0.1 nM) and cause larva mortility (70% of mortality) [32], toxicity against hep G2 (ED50 of 7.4 ± 0.1 μg/mL), lung A549 (ED50 of 149 ± 7 μg/mL) and MCF-7 cell cancer (ED50 of 17 ± 2 μg/mL) [46]. |
| 33. Gigantetrocin, gigantetrocin A, gigantetrocin B [49] | Seeds, leaves | Cytotoxic [49] | Gigantetrocin A and gigantetrocin B exert toxicity against colon cancer cells HT-29, ED50 of 1.24 and 4.1 × 10−5 μg/mL respectively and against lung A549, 3.48 × 10−3 and 2.5 × 10−1 μg/mL respectively [49]. |
| 34. Cis-goniothalamicin [40] | Seeds | Cytotoxic [40] | Crown gall cancer cell inhibition (47%), brine shrimp toxicity (LC50 of 5.2 μg/mL), lung A549 (IC50 of 1.3 × 10−1 μg/mL), breast MCF-7 (1.05 μg/mL), and colon HT-29 cancer cell toxicity (5.3 × 10−3 μg/mL) [40]. |
| 35. (2,4-cis and -trans) gigantetrocinone [57] | Seeds | Cytotoxic [57] | Toxicity on A549 cell lines (ED50 9.73 × 10−3 μg/mL), breast MCF-7 (ED50 2.74 × 10−2 μg/mL) and colon HT-29 (ED50 5.49 × 10−4 μg/mL) [57]. |
| 36. Isoannonacin [49] | Seeds | Cytotoxic [49] | Toxicity against colon HT-29 (ED50 2 × 10−3 μg/mL) and lung tumor A549 (ED50 2 × 10−2 μg/mL) [49]. |
| 37. (2,4-cis and -trans)-isoannonacin [57] | Seeds, leaves | Cytotoxic [57] | Toxicity against breast MCF-7 (ED50 of <10−3 μg/mL), lung A-549 (ED50 of 4.42 × 10−5 μg/mL), and colon HT-29 cell line (ED50 of 1.70 × 10−1 μg/mL) [57]. |
| 38. Isoannonacin-10-one [58] | Seeds | Cytotoxic [58] | Toxicity against colon HT-29 cell line (ED50 of 9x10−3 μg/mL) and lung A549 (ED50 of 7x10−2 μg/mL) [58]. |
| 39. Javoricin [40] | Seeds | Cytotoxic [40] | Toxicity against brine shrimp (LC50 of 4.9 μg/mL), lung A549 (1.7 × 10−2 μg/mL), breast MCF-7 (2.3 × 10−1 μg/mL), and colon HT-29 cancer cells (1.8 μg/mL), as well as inhibition of crown gall tumours (47%) [40]. |
| 40. Longifolicin [59] | Seeds | Cytotoxic [59] | Toxicity in the presence of human hepatoma cells, hep G2 (IC50 = 4.04 μg/mL) and hep 2,2,15 (IC50 = 4.90 × 10−3 μg/mL) [59] |
| 41. Muricapentocin [35] | Leaves | Cytotoxic [35] | Toxicity towards pancreatic carcinoma PACA-2 with ED50 of 5.03 × 10−2 μg/mL and HT-29 colon cancer cells with ED50 value of 7.10 × 10−2 μg/mL [35]. |
| 42. Muricatocin A, B [60] | Leaves | Cytotoxic [60] | Muricatocin A and B showed toxicity against lung cancer cells A549 (ED50 of 7.55 × 10−2 and 3.34 × 10−1 μg/mL), breast MCF-7 (ED50 of 1.23 × 10−1 and 1.03 × 10−1 μg/mL), and colon HT-29 (ED50 of 1.56 and 1.66 μg/mL) [60]. |
| 43. Muricatocin C [61] | Leaves | Cytotoxic [61] | Toxicity against brine shrimp BST (LC50 = 6.04 × 10−1 μg/mL), breast cancer cells MCF-7 (ED50 = 6.45 × 10−2 μg/mL), lung cancer cells A549 (ED50 = 9.09 × 10−2 μg/mL), and colon cancer cells HT-29 (ED50 = 1.48 μg/mL) [61]. |
| 44. Muricin H, I [43] | Seeds | Cytotoxic [43] | Toxicity in the presence of two human hepatoma cells, hep G2 and hep 2,2,15. Muricin H and I showed IC50 of 9.51 × 10−2 and 5.09 × 10−2 μg/mL against hep G2 meanwhile 1.18 × 10−2 and 2.22 × 10−1 μg/mL against hep 2,2,15 [43]. |
| 45. Muricin J, K, L [62] | Fruits | Cytotoxic [62] | Prostate PC-3 cancer cell toxicity [62]. |
| 46. Muricoreacin [63] | Leaves | Cytotoxic [63] | Cytotoxicity on colon HT-29 (ED50 = 0.57 μg/mL), lung A549 (ED50 = 0.23 μg/mL), Breast MCF-7 (ED50 = 1.3 μg/mL), kidney A498 (ED50 = 0.71 μg/mL) and pancreas PACA-2 (ED50 = 2.3 μg/mL) [63]. |
| 47. Murihexocin, murihexocin B, C [32,63] | Leaves, pulp | Cytotoxic [63] | Besides, murihexocin B showed larval mortality of 30% [32]. Then, murihexocin C caused toxicity on colon HT-29 (ED50 = 1.3 μg/mL), lung A549 (ED50 = 1.1 μg/mL), Breast MCF-7 (ED50 = 3.8 μg/mL), kidney A498 (ED50 = 2.5 μg/mL)and pancreas PACA-2 (ED50 = 0.49 μg/mL) [63]. |
| 48. Muricadienin [64] | Roots | Cytotoxic [64] | In vitro inhibitory activity against topoisomerases I and IIα, which are key cell cycle enzymes. Then, high cytotoxicity against HEK293 kidney cancer cells (IC50 = 0.39 µM) [64]. |
| 49. Muridienin 3, 4 [65] | Roots | NR | NR |
| 50. Muricatacin [66] | Seeds | Cytotoxic [66] | Toxicity against lung cancer cells A549 (ED50 = 23.3 μg/mL), colon cancer cells HT-29 (ED50 = 14.0 μg/mL)and breast cancer cells MCF7 (ED50 = 9.8 μg/mL) [66]. |
| 51. Muricatetrocin A, B [59] | Seeds, leaves | Cytotoxic [59] | Toxicity against hepatoma cell line, hep G2 (IC50 = 4.95 × 10−2 μg/mL) and hep 2,2,15 (4.83 × 10−3 μg/mL) [59]. |
| 52. Muricenin [67] | Pulp | Cytotoxic | Toxicity effect against PC-3 cell [67]. |
| 53. Murihexol [68] | Seeds | NR | NR |
| 54. Murisolin [54] | Seeds | Cytotoxic [54] | Toxicity against oral cancer KB cell (ED50 of 1 × 10−2 μg/mL) and VERO cells (ED50 of 1 × 10−1) [54]. |
| 55. Cis-reticulatacin [69] | Roots | Anti-protozoal, anti-lymphoma [69] | Inhibition against Giardia lamblia (IC50 = 59.6 μg/mL) and Entamoeba histolytica (IC50 = 36.5 μg/mL), anti-lymphoma effect against human lymphoma U937 cell line (EC50 = 4.9 ± 0.10 mg/kg) [69]. |
| 56. Rolliniastatin 1 [46] | Seeds | Cytotoxic [46] | Toxicity to lung A549 (ED50 of >500 μg/mL), hepatoma hep G2 (ED50 of 12 ± 2 μg/mL), colon HT-29 (ED50 of 160 ± 36 μg/mL) and breast MCF-7 (ED50 of 17 ± 2 μg/mL) [46]. |
| 57. Solamin [54] | Leaves, seeds. roots | Cytotoxic [54] | Toxicity to oral KB cancer cells (3 × 10−1 μg/mL) and kidney VERO cells (ED50 1 μg/mL) [54]. |
| 58. Squamocin [44] | Seeds | Cytotoxic [44] | Toxicity against hepatoma hep G2 (5.47 × 10−4 μg/mL) and hep 2,2,15 (9.23 × 10−4 μg/mL) [44]. |
| 59. Cis-squamostatin A, squamostatin D [69] | Seeds | Cytotoxic [69] | Cis-squamostatin A and squamostatin D exert toxicity on taxol (resistant cell line) with IC50 of 17.40 ± 2.09 μg/mL and 16.19 ± 1.98 μg/mL respectively [69]. |
| 60. Xylomaticin [69] | Seeds | Cytotoxic [69] | NR |
| Alkaloids | |||
| 1. Anonaine [70] | Leaves, fruits | Neurotoxic, anti-depressive, anti-plasmodium, dopamine inhibitor, cytotoxic [70] | NR |
| 2. Annonaine [71] | Fruits | Anti-depressive [71] | NR |
| 3. Annonamine [72] | Leaves | Neurotoxicity [72] | Toxicity against neuroblastoma SH-SY5Y (IC50 = 195.8 ± 17.0 µM) [72] |
| 4. Asimilobine [71] | Fruits, leaves | Anti-depressive, cytotoxic [71] | NR |
| 5. Coreximine [73] | Leaves, barks, roots, stems | Neurotoxic [73] | NR |
| 6. Isoboldine [74] | Leaves | Anti-malarial [74] | NR |
| 7. Isolaureline [70] | Leaves | Cytotoxic [70] | NR |
| 8. Nornuciferine [71] | Fruits | Anti-depressive [71] | NR |
| 9. (S)-norcorydine [72] | Leaves | Cytotoxic [72] | Toxicity against neuroblastoma SH-SY5Y (IC50 = 186.6 ± 29.4 µM) [72] |
| 10. Reticuline [73] | Leaves, barks, roots, stems | Neurotoxic [73] | NR |
| 11. Swainsonine [75] | Leaves, stems | Immune response stimulation [75] | Inhibit lysosomal acidic α-mannosidase to cause neurotoxicity [75] |
| Phenolic Compounds | |||
| 1. Apigenin-6-C-glucoside [76] | Leaves | Antioxidant [76] | NR |
| 2. Argentinine [77] | Leaves | Antioxidant [77] | NR |
| 3. Caffeoylquinic acid [78] | Leaves, pulps | NR | NR |
| 4. Catechin [77] | Leaves | Antioxidant [77] | NR |
| 5. Coumaric acid [79] | Fruits | NR | NR |
| 6. Daidzein [79] | Leaves | NR | NR |
| 7. Emodin [79] | Leaves | NR | NR |
| 8. Epicatechin [77] | Leaves | NR | NR |
| 9. Gallic acid [77] | Leaves | NR | NR |
| 10. Gallocatechin | Leaves | NR | NR |
| 11. Homoorientin [79] | Leaves | Antioxidant [79] | NR |
| 12. Kaempferol [77] | Leaves, pulp | Antioxidant [77] | NR |
| 13. Kaempferol 3-O-rutinoside [77] | Leaves, pulp | Antioxidant [77] | NR |
| 14. Robinetin [76] | Leaves | Antioxidant [76] | NR |
| 15. Quercetin [77] | Leaves | Antioxidant [77] | NR |
| 16. Quercetin 3-O-glucoside [77] | Leaves | Antioxidant [77] | NR |
| 17. Quercetin 3-O-neohesperidoside [77] | Leaves | Antioxidant [77] | NR |
| 18. Quercetin 3-O-robinoside [77] | Leaves | Antioxidant [77] | NR |
| 19. Quercetin 3-O-rutinoside (Rutin) [77] | Leaves | Antioxidant [77] | NR |
| Part of A. muricata Extract Used/Test Model/Duration | Dose-Route Administration | Findings of Study | Possible Metabolites | References |
|---|---|---|---|---|
| • Ethanolic- water (leaves) • C57BL/6J male mice, T2DM (Intraperitoneal (i.p.) injection of streptozotocin (STZ; 60 mg/kg) at once a week for a second consecutive week) • Daily for 9 weeks | Orally gavaged: Group 1: Normal control, NC Group 2: Diabetes mellitus control, DMC Group 3: Low dosage of AME treated group (LAM); DMC + 50 mg/kg of extract Group 4: High dosage of extract treated group (HAM); DMC + 100 mg/kg of extract | (a) Body weight: • No changes between extract treated group and DMC groups (b) Liver Weight: • Significant changes between NC and DMC with extract treated rats (c) Diet Intake: • No changes among all groups (d) FBG and HbA1c: • Rats in LAM & HAM significantly had lower FBG and HbA1c levels compared to the DMC group (e) Glucose intolerance and insulin resistance • Significant difference in blood-glucose levels between extract treated and DMC rats at 30 and 60 min after oral glucose administration (f) Hepatic damage: • Rats in the LAM group has significantly reduced ALT level when compared to DMC, but not for HAM group (g) Hepatic Morphology and Hepatic Triglyceride (TG) and Total Cholesterol (TC): • Lipid droplets and their size were decreased in the AME rats compared to DMC • Hepatic TG reduced to almost normal in LAM rats compared to DMC • No changes for hepatic TC content (h) Insulin Signaling: • Significant difference in IRS-1 and GLUT2 proteins between LAM and the DMC groups (i) Hepatic Oxidative Stress: • A significant difference in levels of 4-HNE, protein carbonyls, Nrf2 and NQO1 between the extract treated rats and DMC (j) Energy Metabolism: • LAM-treated rats have normalized energy metabolism through increased levels of p-AMPK and PGC1α. • p-mTOR was significantly decreased in the treatment groups compared to DMC. (k) Autophagy: • The LC3-II expression level was significantly increased in extract treated groups compared to DMC (l) Lipid metabolism • The proteins related to fat synthesis in LAM treated rats were significantly reduced to normal levels compared to DMC | • Total phenolic • Acetogenins, rutin, quercetin, and kaempferol • Rutin, kaempferol-3-O-rutinoside, quercetin, kaempferol, muricoreacin, annonacin, and annonacinone | [89] |
| • Ethanol extracts (leaves) • 5 groups; 2 control groups and 3 treatment groups [Male white rats (Rattus norvegicus). Diabetic rats (STZ-intraperitoneally induced)] • 1 week | (a) Group 1—control (b) Group 2—Acarbose (1.8 mg/kg) (c) Group 3—10 mg/kg (d) Group 3—20 mg/kg (e) Group 3—30 mg/kg | Pre-test: (a) No changes in blood glucose for the pre-test experimental groups Post-test: (a) No changes for the negative control group p > 0.05 (b) All treatment groups and the positive control group obtained p < 0.05, which means showing significant changes in blood glucose (reduction) The highest anti-hyperglycemic effect was treatment group 3 (30 mg/kg of soursop leaf extract), and it was similar to acarbose group in reducing blood glucose | • Flavonoids (quercetin and chrysin), • Alkaloids • Tannins • Ellagic acid | [90] |
| • Ethanol extracts (leaves) • 5 groups; 2 control groups and 3 treatment groups [Wistar rats, diabetic rats (alloxan-intraperitoneally induced)]. • 21 days of treatment | Orally gavaged: (a) Group 1—control (b) Group 2—Alloxan treated (c) Group 3—50 mg/kg of extract (d) Group 3—100 mg/kg of extract (e) Group 3—200 mg/kg of extract | • Rats in groups 2 to 5 displayed hyperglycemia after alloxan inducement • Significantly reduce blood glucose for groups treated with the extract • Rats in groups 2 to 5 displayed hypercholesterolemia after alloxan inducement • Significantly reduce hypercholesterolemia for groups treated with the extract • No changes in islet of Langerhans | • Flavonoids • Saponins • Phytosterols • Phenolics | [91] |
| • Ethanol extracts (fruit) • Groups: 1. α-Amylase Inhibition Assay 2. α-Glucosidase Inhibition Assay 3. Angiotensin-I Converting Enzyme (ACE) Inhibition Assay 4. Free Radical Scavenging Ability 5. Total Antioxidant Power 6. Hydroxyl (OH) Radical Scavenging Assay 7. Determination of Reducing Property 8. Fe2+ Chelation Assay 9. Determination of Total Phenol Content 10. Determination of Total Flavonoid Content | In vitro | 1. α-Amylase and α-Glucosidase Inhibition Assays: • Extracts inhibited α- amylase, α-Glucosidase and ACE activities in a concentration-dependent manner • The highest inhibition: pericarp extract • The lowest inhibition: seed extract • Extracts showed a lower inhibitory effect when compared to acarbose and captopril in α-amylase, α-glucosidase, and ACE inhibition assay 2. All extracts scavenged DPPH radical in a concentration-dependent manner • The highest DPPH radical scavenging ability: pericarp extract 3. The pericarp extract had the highest scavenging ability of the ABTS radical cation 4. All extracts significantly scavenged OH radical in concentration-dependent manner • The pericarp extract had the highest scavenging ability of the OH free radical • The lowest inhibition: seed extract 5. Extracts chelated Fe2+ in concentration-dependent manner • The highest Fe2+ chelating ability: pericarp extract • The lowest chelating ability: the seed 6. Pericarp extract had the highest total phenol and flavonoid content followed by the pulp and the seed. | • Phenolics | [5] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zubaidi, S.N.; Mohd Nani, H.; Ahmad Kamal, M.S.; Abdul Qayyum, T.; Maarof, S.; Afzan, A.; Mohmad Misnan, N.; Hamezah, H.S.; Baharum, S.N.; Mediani, A. Annona muricata: Comprehensive Review on the Ethnomedicinal, Phytochemistry, and Pharmacological Aspects Focusing on Antidiabetic Properties. Life 2023, 13, 353. https://doi.org/10.3390/life13020353
Zubaidi SN, Mohd Nani H, Ahmad Kamal MS, Abdul Qayyum T, Maarof S, Afzan A, Mohmad Misnan N, Hamezah HS, Baharum SN, Mediani A. Annona muricata: Comprehensive Review on the Ethnomedicinal, Phytochemistry, and Pharmacological Aspects Focusing on Antidiabetic Properties. Life. 2023; 13(2):353. https://doi.org/10.3390/life13020353
Chicago/Turabian StyleZubaidi, Siti Norliyana, Hidayah Mohd Nani, Mohd Saleh Ahmad Kamal, Taha Abdul Qayyum, Syahida Maarof, Adlin Afzan, Norazlan Mohmad Misnan, Hamizah Shahirah Hamezah, Syarul Nataqain Baharum, and Ahmed Mediani. 2023. "Annona muricata: Comprehensive Review on the Ethnomedicinal, Phytochemistry, and Pharmacological Aspects Focusing on Antidiabetic Properties" Life 13, no. 2: 353. https://doi.org/10.3390/life13020353