
First Fossil Record of Trichomanes sensu lato (Hymenophyllaceae) from the Mid-Cretaceous Kachin Amber, Myanmar


1
State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China
2
Department of Botany, National Museum of Nature and Science, 4-1-1 Amakubo, Tsukuba 305-0005, Japan
3
Komarov Botanical Institute of the Russian Academy of Sciences, Prof. Popova Str. 2, Saint Petersburg 197376, Russia
4
College of Agro-Grassland Science, Nanjing Agricultural University, Nanjing 210095, China
5
Lushan Botanical Garden, Chinese Academy of Sciences, Jiujiang 332900, China
*
Authors to whom correspondence should be addressed.
Life 2023, 13(8), 1709; https://doi.org/10.3390/life13081709
Submission received: 7 June 2023
/
Revised: 4 August 2023
/
Accepted: 7 August 2023
/
Published: 9 August 2023
(This article belongs to the Special Issue Recent Research on Palaeontology)
Abstract
:Hymenophyllaceae (filmy ferns), with ca. 430 species, are the most species-rich family of early diverging leptosporangiate ferns but have a poor fossil record dating back to the Late Triassic period. Traditionally, Hymenophyllaceae comprise two species-rich genera or clades: Hymenophyllum (hymenophylloids) and Trichomanes sensu lato (s.l.) (trichomanoids). Unequivocal fossils of Hymenophyllum have been reported from the Early Cretaceous of central Mongolia and the early Eocene of Okanogan Highlands, Washington, USA. However, despite being a highly diversified lineage with an estimated 184 extant species, Trichomanes s.l. lack a definitive fossil record, which severely affects the reliability of the molecular dating of this group. Here, we report the first unequivocal fossil record of Trichomanes s.l. as T. angustum comb. nov. on the basis of fertile material with tubular involucres and long exserted receptacles from the mid-Cretaceous Kachin amber, Myanmar. This species was previously tentatively assigned to Hymenophyllites due to a lack of fertile evidence. Inferred to be an epiphytic fern, T. angustum further enriches the species diversity of the epiphytic palaeocommunities in the mid-Cretaceous Kachin amber, which are mainly composed of Porellalean leafy liverworts and Dicranalean and Hypnodendralean mosses. Fossil records indicate that Hymenophyllaceae probably originated in the tropical Pangea at the latest in the Triassic when all continents were coalesced into a single landmass and had already accumulated some notable diversity in low-middle latitude areas of Laurasia by the mid-Cretaceous period.
1. Introduction
Hymenophyllaceae (filmy ferns) include ca. 430 extant species [1] and are the largest family of early diverging leptosporangiate ferns [1,2,3,4,5,6,7,8]. Hymenophyllaceae are predominantly distributed in moist mossy forests in tropical mountains and south-temperate areas [9] and are mainly composed of epiphytic ferns but also include some terrestrial and climbing species [10,11]. Hymenophyllaceae are mainly characterized by a reduction of the leaf lamina into usually one-cell-layer thick lamina between the veins with the absence of stomata, and the presence of marginal indusiate sori, sporangia with an oblique annulus, and trilete spores [4,9,12,13,14,15,16]. The phylogenetic position of Hymenophyllaceae was not well resolved [1,4] until 2018. The phylogenomic analyses using transcriptome sequencing data strongly support the idea that Hymenophyllaceae and Gleicheniaceae formed a monophyletic clade sister to Dipteridaceae [7,8]. This phylogenetic relationship was also supported by sporangia characters [8]. Traditionally, Hymenophyllaceae comprise two species-rich genera or clades: Hymenophyllum Sm. (hymenophylloid) and Trichomanes L. sensu lato (s.l.) (trichomanoid) [13,16,17,18,19]. Hymenophyllum has bivalvate involucres (indusia) with receptacles usually included within the valves or rarely and shortly exserted, while Trichomanes s.l. is characterized morphologically by its tubular or campanulate involucres and hair-like receptacles that are usually long-exserted [16].
Although it is clear that filmy ferns are ancient lineages according to their near-basal phylogenetic position in early-diverging leptosporangiate ferns [1,5,6,7,8,20], reliable fossils of this family are scarce due to low fossilization potential of the delicate, membranous laminae [9,11,21,22,23,24], hampering detailed reconstructions of the origins of their extant diversity. The earliest convincing fossils of the Hymenophyllaceae were reported as Hopetedia praetermissa Axsmith et al. from the Late Triassic (Carnian) Pekin Formation of North Carolina, USA [21]. Hopetedia Axsmith et al. display a character state mosaic of the two extant genera, namely Trichomanes-like funnel-shaped indusia and Hymenophyllum-like included receptacles [21]. Eogonocormus Deng from the Lower Cretaceous of northeastern China is a small thalloid plant with creeping rhizomes and marginal sori with in situ spores, borne on fanlike pinnule lobes, which indicate convincingly that this genus belongs to the Hymenophyllaceae [25]. Reliable filmy fern fossils also include Hymenophyllites H.R.Goeppert from the mid-Cretaceous of Kazakhstan and Myanmar [26,27] and Acrostichopteris Fontaine from the Early Cretaceous of Spain, China and USA [28,29,30,31]. However, it is uncertain with respect to the affinities between the above-mentioned fossil genera and the extant genera of Hymenophyllaceae. The earliest fossils of Hymenophyllum have been described as H. iwatsukii Herrera et al. based on abundant and exceptionally well-preserved lignified material from the Early Cretaceous (Aptian–Albian) of Mongolia [11]. Hymenophyllum axsmithii Pigg et al. was described from the early Eocene Okanogan Highlands, Washington, USA [24]. However, despite being a highly diversified lineage with an estimated 184 extant species [1], Trichomanes s.l. lack a definitive fossil record to calibrate its internal nodes, which severely affects the reliability and the dating of this group of ferns.
Recently, Li et al. [27] described some sterile lamina fragments from the mid-Cretaceous Kachin amber, Myanmar, as three new fossil species of Hymenophyllites, including H. angustus Y.Li et Y.-D.Wang, H. kachinensis Y.Li et Y.-D.Wang, and H. setosus Y.Li et Y.-D.Wang. In the present paper, we transfer H. angustus to Trichomanes s.l. based on newly found fertile material from Kachin amber, Myanmar. Trichomanes angustum comb. nov. displays typical trichomanoid characters, namely tubular involucres and long exserted receptacles, and thus represents the first unequivocal fossil trichomanoid filmy fern known so far. We also discuss its palaeoecological implications and infer the palaeogeographic history of Hymenophyllaceae based on fossil records.
2. Materials and Methods
Kachin amber originates from several amber mines about 20 km southwest of the village of Tanai in the Hukuang Valley of Kachin State, northern Myanmar [32,33]. Kachin amber deposits are currently the most important source of Cretaceous amber-preserved paleobiota [32,34] and yield a large number of plant and animal inclusions [35,36]. The age of Kachin amber is regarded as the late Albian–early Cenomanian, based on the evidence of the ammonite Puzosia Matsumoto and palynomorphs [37,38]. The U-Pb dating of zircons suggests the earliest Cenomanian age (98.79 ± 0.62 Ma) for the amber-bearing horizon of Kachin amber [34].
The fossils studied here include two pieces of Kachin amber from Myanmar and are housed at the Collection Department of Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China, under accession numbers PB200744 and PB201715. Concerning the recent conflicts in Myanmar [39], we declare that we followed the recommendations by Haug et al. [40]. All Kachin amber pieces mentioned in this study were acquired in compliance with the laws of Myanmar and China, including Myanmar’s import and export regulations of jewelry and China’s fossil law. The extant specimens were deposited at the National Museum of Nature and Science, Tsukuba, Japan.
The amber inclusions were studied under a ZEISS Axio Zoom.V16 microscope (Carl Zeiss AG, Oberkochen, Germany) equipped with a high-resolution digital camera (Axiocam 512 color, Carl Zeiss AG, Oberkochen, Germany). Incident and transmitted lights were used simultaneously for photography. All images were digitally stacked photomicrographic composites of ca. 20–50 individual focal planes using the software package ZEN 2.3 pro for a better illustration of the three-dimensional inclusions. The palaeocoordinates of fossil sites were converted from their extant coordinates using PointTracker v4c software and plotted on individual palaeogeographic maps (Mollweide projection) at three time points, i.e., Late Triassic (220 Ma), mid-Cretaceous (120 Ma), and early Eocene (50 Ma), using ArcView GIS 3.2 software. The terminology for the description of the present fern fossils follows Axsmith et al. [21] and Liu et al. [14].
3. Results
Systematic Paleontology
Order: Hymenophyllales A.B.Frank
Family: Hymenophyllaceae Mart.
Genus: Trichomanes L. sensu lato
Species: Trichomanes angustum (Y.Li et Y.-D.Wang) Y.Li et Ebihara, comb. nov.
Synonym: Hymenophyllites angustus Y.Li et Y.-D.Wang [27] (pp. 3–5, Figure 1)
Holotype and Paratypes: PB200744a and PB200744b–h
New specimens: PB201715a, b
Age: Late Albian–early Cenomanian, mid-Cretaceous.
Type locality: Amber mines southwest of the village of Tanai ca. 105 km north of Myitkyina in Kachin State, northern Myanmar.
Repository: Collection Department of Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences, Nanjing, China.
Specific diagnosis: Leaf fragments tripinnate, glabrous. Pinnae leaf fragments closely spaced and alternate. Pinnules of leaf fragments are closely spaced, anadromous, distinctly divided into simple or forked segments. Segments dichotomized 0–3 times to form one to several ultimate lobes. Lobes narrow, slightly elongated, (0.1–) 0.3–0.4 mm wide, entire-margined, with a single veinlet. Lamina venation is anadromous without false veinlets. Differentiated marginal elongated cells are absent. Sori paratactic, borne at the apex of the first arising lobes of the pinnule or its segments. Involucres tubular, ca. 3–5 times as long as wide. Involucre mouths are usually non-dilated. Receptacles are filiform and long exserted.
Description: The two large leaf fragments (PB200744a and PB201715b) are likely lamina portions, tripinnate, and are up to ca. 2.1 cm long (Figure 1A,B and Figure 2A,B). However, it is also possible that they actually stem from single large lateral pinnae, as stripes have not been found yet. Lamina is membranaceous, one cell layer thick between the veins, and glabrous (Figure 1C–F and Figure 2C,D). The lamina rachis (also possibly pinna costa) is not winged or winged only at the apex, glabrous (Figure 1B and Figure 2A,B). The pinnae of leaf fragments are at least in 3 pairs, closely spaced, alternate, spread, triangular-ovate, up to 1.2 cm long (Figure 1A,B and Figure 2A,B). The pinnules of leaf fragments are closely spaced, anadromous, up to 0.8 cm long, often with the most proximal ones overlapping the rachis (Figure 1A,B and Figure 2A,B). The pinnules are distinctly divided into segments which are further dichotomized 0–3 times to form simple to several ultimate lobes (Figure 1A,B and Figure 2A–C). The lobes are flat, narrow, slightly elongated, (0.1–) 0.3–0.4 mm wide, entire-margined, with acute, obtuse, truncate to retuse apices (Figure 1C–E, Figure 2C, Figure 3A,B and Figure 4). Each lobe is vascularized by a single veinlet. Lamina venation is anadromous and dichotomous to form more or less zigzag costae and costules. False veinlets are absent. Laminar cells are more than 3 rows between the midrib and margins, polygonal, isodiametrical to slightly elongated with thin and straight cell walls, 25–88 μm long and 14–66 μm wide outside the venation area; however, they are also elongated fusiform to rectangular inside the venation area (Figure 1F and Figure 2D). Differentiated marginal elongated cells are absent. Stomata are absent. Sori are paratactic, borne at the apex of the first arising lobes of the pinnule or its segments, oriented upward, nearly straight to the plane of the lamina (Figure 2A,B and Figure 3A–E). Involucres are tubular, slightly curved, immersed in lobes or segments, winged throughout, 1.0–2.2 mm long and 0.3–0.6 mm wide, ca. 3–5 times as long as wide (Figure 3 and Figure 4). Involucre mouths are usually not dilated (Figure 3C,E–G and Figure 4). Receptacles are filiform, long exserted, and up to 3.1 mm long (Figure 3B,C,F,G and Figure 4).
Remarks: Although Ebihara et al. [3] subdivided the Trichomanes s.l. into eight genera mainly based on the results of molecular phylogeny [3,13], it was virtually impossible to find macro-morphological characters that consistently discriminated these genera for all species [16]. It is especially difficult to put this classification into practice for the present fossils because they apparently lack their rhizome part: a very important character for generic identification. So, we adopted the concept of Trichomanes s.l. [17,18,19], which is still used today [16,41].
It is strange that some pollen grains adhere on the lamina surface in PB201715 (Figure 5A,B), where filmy fern sporangia and spores are completely absent, although there are rich sori and involucres. These pollen grains are monoporate, with a round to rounded triangle, 31–65 μm in diameter (Figure 5C–F). Monoporate pollen is indeed present in some Mesozoic gymnosperm taxa, such as Admolia Batten and Perinopollenites Couper [42]. The former is an unknown gymnosperm reported from the Early Cretaceous of the UK and China, while the latter belongs to taxodioid Cupressaceae (‘Taxodiaceae’) and is widely distributed around the world during the Jurassic to the Cretaceous period [42]. However, the lack of ornamentation details hampers a definite identification.
4. Discussion
4.1. Comparisons
Although highly challenging, we made a great effort to narrow down this identification according to the classification of Ebihara et al. [3], who subdivided Trichomanes s.l. into eight genera, namely Abrodictyum C.Presl, Callistopteris Copel., Cephalomanes C.Presl, Crepidomanes C.Presl, Didymoglossum Desv., Polyphlebium Copel., Trichomanes L. sensu stricto (s.str.), and Vandenboschia Copel., each with one to four subgenera. Firstly, our fern remains could be distinguished from the Palaeotropical genera Callistopteris and Cephalomanes as well as the predominantly Neotropical genus Trichomanes s.str. by having tubular involucres, while the latter three genera had campanulate involucres [3,43]. In addition, our remains were divided at least tripinnate with one cell layer thick pinnae as well as anadromous venation, while Cephalomanes only has a once-pinnate lamina with asymmetric pinnae and Trichomanes s.str. also displayed other different features in its five subgenera. For example, once-pinnate to bi-pinnate-pinatifid lamina was found in subgenera Afrotrichomanes and Lacostea, more than one cell layer thick lamina was identified in subgenera Davalliopsis and Feea, and usually, catadromous venation is found in subgenus Trichomanes [3]. Secondly, our fern remains differ from Abrodictyum in having more than three rows of laminar cells between the midrib and margins and having straight internal cell walls, while the lamina of most Abrodictyum species is reduced to less than three rows of cells between the midrib and the margins and internal cell walls are wavy or pitted [3,14,44]. Finally, our fern remains could be confined to a hemiepiphytic/epiphytic clade within the trichomanoids, comprising Polyphlebium, Didymoglossum, Crepidomanes, and Vandenboschia [10]; however, the absence of false veinlets clearly differentiated our fern remains from Didymoglossum that always have false veinlets [3,14]. Therefore, we could only narrow down the identification of our fossils to three candidate genera, namely Polyphlebium, Crepidomanes, and Vandenboschia. Unfortunately, we have not found any other traits in our fossils that allow the clear-cut identification of these three genera by lamina characters alone (Figure 6). To identify the genus, we needed to add more evidence, such as rhizome and rachis characters.
Polyphlebium is a primarily pantropical genus with ca. 15 species, mainly distributed in the Southern Hemisphere [3,45,46,47,48]. The present fossils had more divided lamina than most species of Polyphlebium, except the two strictly Neotropical species, P. angustatum (Carmich.) Ebihara et Dubuisson (Figure 6A) and P. capillaceum (L.) Ebihara et Dubuisson, one endemic species, P. exsectum (Kunze) Ebihara et Dubuisson, in Chile, as well as a Central and South American species P. hymenophylloides (Bosch) Ebihara et Dubuisson. However, P. capillaceum has filiform linear ultimate lobes, and P. hymenophylloides has differentiated marginal elongate cells [48,49]. The present fossils look very similar to P. angustatum in gross morphology [48] (Figure 6A) but still differ from it in having extremely narrow ultimate lobes (usually 0.3–0.4 mm wide) and non-dilated involucre mouths. The ultimate lobes in P. angustatum are 0.5–0.7 wide [48].
Crepidomanes include two subgenera, Crepidomanes and Nesopteris, and ca. 30 species, distributed from Palaeotropics to northern temperate regions [3]. In addition, one species in the Neotropics (C. pyxidiferum (L.) Dubuisson et Ebihara) was found [50]. The subgenus Crepidomanes includes three sections and differs from our fern remains as follows. In section Crepidomanes, false veinlets are often present, and involucre mouths are usually bilabiate; section Gonocormus has campanulate involucre with dilate mouths; section Crepidium has double rows of elongate marginal cells [3]. It is to be noted that some members of section Crepidomanes lack false veinlets, e.g., C. schmidtianum (Zenker ex Taschner) K.Iwats. (Figure 6C) and C. vitiense (Baker) Bostock. The subgenus Nesopteris has rather large and more than 15 cm long fronds [3], but the relatively long involucres in our fossils reminded us of this subgenus (Figure 6D).
Vandenboschia is a pantropical genus with two subgenera, Vandenboschia and Lacosteopsis, and ca. 15 species that are often hemiepiphytic and occasionally terrestrial [3,10,46]. The subgenus Vandenboschia owns bipinnate or more finely divided lamina (Figure 6B), while the subgenus Lacosteopsis has once pinnate lamina [3]. Thus, the identification of the subgenus Lacosteopsis could be excluded. In addition, this genus always has minute clavate hairs on its rachises [3]. If our two large fossils (Figure 1A,B and Figure 2A,B) stem from the lamina portion rather than from single large lateral pinnae, the identification of subgenus Vandenboschia could also be excluded owing to its minute clavate hairs on rachises.
4.2. Palaeoecological Implications
Epiphytes are non-parasitic plants that germinate and grow on phorophytes at all stages of life [51]. Thus, epiphytes and their host phorophytes form a typical commensal relationship. Filmy ferns represent mostly a dense community of low epiphytes in the forest understory; however, some species can fully cover trunks and branches in cloud forests [44,52,53]. It is probable that the fossil species Trichomanes angustum was an epiphyte because it could be morphologically confined to a hemiepiphytic/epiphytic clade within the trichomanoids [10]. According to the analysis of two-gene (rbcL + rps4), the crown group of this hemiepiphytic/epiphytic clade started to evolve around 121.7 ± 12.3 Ma [10], which is in agreement with our assignation of T. angustum to this clade. The mid-Cretaceous Kachin amber forest was highly tropical and probably located close to the seashore [32,37,38]. It was rich in epiphytes that are known to be mainly composed of Porellalean leafy liverworts [54,55,56,57,58,59,60,61,62,63,64,65] and Dicranalean and Hypnodendralean mosses [66,67,68,69,70]. The discovery of T. angustum, along with Hymenophyllites kachinensis and H. setosus in the mid-Cretaceous Kachin amber [27], further enriches the species diversity of the epiphytic palaeocommunities.
There are still some intriguing aspects of T. angustum that remain unknown, including the host phorophytes. However, it is likely that T. angustum grew on resin-producing gymnosperm trees so that it was easily captured by the resin, and the monoporate pollen adhering on the fern lamina surface (Figure 5) represents pollen rain from the phorophyte trees or nearby trees. The mid-Cretaceous Kachin amber forest was dominated by various gymnosperm trees, including members of Araucariaceae and Cupressaceae [32,37]. Nuclear magnetic resonance spectroscopic studies and anatomical analyses of fossil wood fibers have indicated that araucarioid trees of the Araucariaceae, especially Agathis Salisb., were determined as the source of Kachin amber [71].
4.3. Paleobiogeographic History of Hymenophyllaceae
Copeland [72] suggested an Antarctic origin for the Hymenophyllaceae according to the fact that most filmy ferns grow in the Southern Hemisphere, with numerous monotypic groups in austral regions. Iwatsuki [9,73] believed that filmy ferns evolved in the tropics and subsequently dispersed from there. From the analysis of rbcL sequence data, Dubuisson et al. [74] inferred a basal position for Asian groups in Trichomanes, while the most basal taxa within Hymenophyllum are also Paleotropical and/or austral [75]. Thus, Dubuisson et al. [74] speculated that Hymenophyllaceae probably arose and first diverged in the Paleotropics, possibly in Asia. Divergence time estimates indicate that Hymenophyllaceae might have evolved during the Carboniferous to the Triassic period [2,5,6,10,20,43] and split into hymenophylloids and trichomanoids in the Middle Jurassic period [10]. Numerous Paleozoic and Mesozoic Hymenophyllaceae-like fronds were assigned to the extinct genera Hymenophyllites, Trichomanides, and Trichomanites [76,77,78,79] or to the extant genus Hymenophyllum [80], but the affinity of these fossils to Hymenophyllaceae is quite uncertain owing to the lack of definite evidence of a membranaceous habit or marginal indusiate sori [21,81].
The earliest reliable Hymenophyllaceae fossils are reported from the Late Triassic Pekin Formation of North Carolina, USA [21], followed by an apparent gap in the Jurassic period (Table 1; Figure 7). Hymenophyllaceae diversified in the Early to middle Cretaceous period of Laurasia with several fossil genera and species reported from Spain, Kazakhstan, Mongolia, China, and the USA [11,25,26,27,28,29,30,31] (Table 1; Figure 7). Some younger fossils of Hymenophyllaceae have been recently reported to be from the early Eocene of Okanogan Highlands, Washington, USA [24] (Table 1; Figure 7). It is strange that all unequivocal fossils of Hymenophyllaceae were reported from the Northern Hemisphere. Despite being a worldwide fern lineage predominantly distributed in tropical mountains and south-temperate areas, Hymenophyllaceae lack a definitive fossil record in the Southern Hemisphere (Figure 7). Although some putative fossils of Hymenophyllaceae were reported as Trichomanides laxum Tenison-Woods and T. spinifolium Tenison-Woods from the Jurassic of Queensland, Australia [76] and Hymenophyllum priscum Menéndez from the Late Cretaceous of Chile [80], the identification to Hymenophyllaceae has been thought to be wrong or less convincing [21]. Although the bulk of the extant diversity of Hymenophyllaceae appears to have accumulated later in angiosperm-dominated forests during the Cenozoic [10], the reliable fossil record of Hymenophyllaceae is scarce in the Cenozoic (Table 1; Figure 7).
All in all, present fossil evidence indicates a tropical Pangea origin for Hymenophyllaceae at the latest in the Triassic when all continents coalesced into a single landmass Pangea under a global hothouse climate [82] (Figure 7), and some notable diversity was already developed in low and middle latitude areas of Laurasia in the mid-Cretaceous when the earth experienced a hothouse climate again [82] (Figure 7).

Table 1.
Reliable fossil record of Hymenophyllaceae.
| Taxon | Age | Stratigraphic Horizon | Locality | Reference |
|---|---|---|---|---|
| Acrostichopteris alcainensis Sender | Middle–late Albian, Early Cretaceous | Upper Member, Escucha Formation | Alcaine village, Teruel Province, Spain | [31] |
| Acrostichopteris fimbriata Knowlton | late Berriasian–Barremian, Early Cretaceous | Kootanie Formation | Meridith mine, Cascade County, Montana, USA | [29] |
| Acrostichopteris interpinnula Meng et Chen | Aptian–Albian, Early Cretaceous | Fuxin Formation | Xinqiu coal mine, Fuxin, Liaoning, China | [30,83] |
| Acrostichopteris longipennis Fontaine | late Aptian–early Albian, Early Cretaceous | Potomac Group | Baltimore, Maryland, and Richmond, Virginia, USA | [28,31] |
| Eogonocormus cretaceum Deng | late Barremian–early Aptian, Early Cretaceous | Lower Coal-bearing Member, Huolinhe Formation | Huolinhe Basin, Inner Mongolia, China | [25,84] |
| Eogonocormus linearifolius Deng | late Barremian–early Aptian, Early Cretaceous | Lower Coal-bearing Member, Huolinhe Formation | Huolinhe Basin, Inner Mongolia, China | [25,84] |
| Hopetedia praetermissa Axsmith et al. | late Carnian, Late Triassic | Middle portion of Pekin Formation | Boren Clay Company pit near Gulf, North Carolina, USA | [21] |
| Hymenophyllites kachinensis Y.Li et Y.-D.Wang | Upper Albian–lower Cenomanian, mid-Cretaceous | No data | Amber mines near Tanai, Kachin State, Myanmar | [27] |
| Hymenophyllites macrosporangiatus Vachrameev | Middle Albian, Early Cretaceous | Kysylshen Formation | Karachetau and Kysylshen, western Kazakhstan | [26] |
| Hymenophyllites setosus Y.Li et Y.-D.Wang | Upper Albian–lower Cenomanian, mid-Cretaceous | No data | Amber mines near Tanai, Kachin State, Myanmar | [27] |
| Hymenophyllum axsmithii Pigg et al. | Ypresian, early Eocene | Tom Thumb Member, Klondike Mountain Formation | Boot Hill, Republic, Washington, USA | [24] |
| Hymenophyllum iwatsukii Herrera et al. | Aptian–Albian, Early Cretaceous | Tevshiin Govi and Khukhteeg Formations | Tevshiin Govi and Tugrug, central Mongolia | [11] |
| Trichomanes angustum comb. nov. | Upper Albian–lower Cenomanian, mid-Cretaceous | No data | Amber mines near Tanai, Kachin State, Myanmar | This paper |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
5. Conclusions
Here, we report the first definitive fossil species of Trichomanes s.l. as T. angustum comb. nov., which is combined from the previously described Hymenophyllites angustus, based on newly found fertile material from the mid-Cretaceous Kachin amber, Myanmar. Morphological comparisons suggest that T. angustum can be confined to a hemiepiphytic/epiphytic clade within the trichomanoid. Inferred to be an epiphytic fern, T. angustum further enriches the species diversity of the epiphytic palaeocommunities in mid-Cretaceous Kachin amber, mainly comprising Porellalean leafy liverworts and Dicranalean and Hypnodendralean mosses. Fossil records indicate that Hymenophyllaceae probably originated in the tropical Pangea at the latest, in the Triassic and accumulated some notable diversity in low and middle-latitude areas of Laurasia by the mid-Cretaceous.
Author Contributions
Conceptualization, Y.L. and Y.-M.C.; methodology, Y.L. and Y.-M.C.; software, Y.L. and Y.-M.C.; validation, Y.L.; formal analysis, Y.L.; investigation, Y.L. and A.E.; resources, Y.L., N.N. and Z.-Z.T.; data curation, Y.L.; writing—original draft preparation, Y.L.; writing—review and editing, Y.L., A.E., N.N., Z.-Z.T. and Y.-M.C.; visualization, Y.L. and A.E.; supervision, Y.L. and Y.-M.C.; project administration, Y.-M.C.; funding acquisition, Y.-M.C. and N.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 42002023, the institutional research project of the BIN RAS (Saint Petersburg, Russia), grant number 122011900029-7 and the State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, CAS, grant number 20192101.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are reported in this paper.
Acknowledgments
We thank the editors for handling our manuscript and appreciate two anonymous reviewers for their valuable suggestions that improved our manuscript. We are also grateful to Jing-Jing Tang for her help in photographing the amber specimens under the ZEISS Axio Zoom.V16 microscope.
Conflicts of Interest
The authors declare no conflict of interest.
References
- The Pteridophyte Phylogeny Group. A community-derived classification for extant lycophytes and ferns. J. Syst. Evol. 2016, 54, 563–603. [Google Scholar] [CrossRef]
- Pryer, K.M.; Schuettpelz, E.; Wolf, P.G.; Schneider, H.; Smith, A.R.; Cranfill, R. Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences. Am. J. Bot. 2004, 91, 1582–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebihara, A.; Dubuisson, J.-Y.; Iwatsuki, K.; Hennequin, S.; Ito, M. A taxonomic revision of Hymenophyllaceae. Blumea 2006, 51, 221–280. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.R.; Pryer, K.M.; Schuettpelz, E.; Korall, P.; Schneider, H.; Wolf, P.G. A classification for extant ferns. Taxon 2006, 55, 705–731. [Google Scholar] [CrossRef]
- Rothfels, C.J.; Li, F.-W.; Sigel, E.M.; Huiet, L.; Larsson, A.; Burge, D.O.; Ruhsam, M.; Deyholos, M.; Soltis, D.E.; Stewart, C.N., Jr.; et al. The evolutionary history of ferns inferred from 25 low-copy nuclear genes. Am. J. Bot. 2015, 102, 1089–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testo, W.; Sundue, M. A 4000-species dataset provides new insight into the evolution of ferns. Mol. Phylogenet. Evol. 2016, 105, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.-P.; Kuo, L.-Y.; Guo, C.; Li, H.; Li, Z.-Y.; Qi, J.; Wang, L.-B.; Hu, Y.; Xiang, J.-Y.; Zhang, C.-F.; et al. A well-resolved fern nuclear phylogeny reveals the evolution history of numerous transcription factor families. Mol. Phylogenet. Evol. 2018, 127, 961–977. [Google Scholar] [CrossRef]
- Shen, H.; Jin, D.-M.; Shu, J.-P.; Zhou, X.-L.; Lei, M.; Wei, R.; Shang, H.; Wei, H.-J.; Zhang, R.; Liu, L.; et al. Large-scale phylogenomic analysis resolves a backbone phylogeny in ferns. GigaScience 2018, 7, gix116. [Google Scholar] [CrossRef] [Green Version]
- Iwatsuki, K. Hymenophyllaceae. In The Families and Genera of Vascular Plants, Vol. 1, Pteridophytes and Gymnosperms; Kramer, K.U., Green, P.S., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1990; pp. 157–163. [Google Scholar]
- Hennequin, S.; Schuettpelz, E.; Pryer, K.M.; Ebihara, A.; Dubuisson, J.-Y. Divergence times and the evolution of epiphytism in filmy ferns (Hymenophyllaceae) revisited. Int. J. Plant Sci. 2008, 169, 1278–1287. [Google Scholar] [CrossRef] [Green Version]
- Herrera, F.; Moran, R.C.; Shi, G.-L.; Ichinnorov, N.; Takahashi, M.; Crane, P.R.; Herendeen, P.S. An exquisitely preserved filmy fern (Hymenophyllaceae) from the Early Cretaceous of Mongolia. Am. J. Bot. 2017, 104, 1370–1381. [Google Scholar] [CrossRef] [Green Version]
- Farrar, D.R. Hymenophyllaceae. In Flora of North America North of Mexico, Vol. 2 Pteridophytes and Gymnosperms; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA; Oxford, UK, 1993; pp. 190–197. [Google Scholar]
- Ebihara, A.; Iwatsuki, K.; Ito, M.; Hennequin, S.; Dubuisson, J.-Y. A global molecular phylogeny of the fern genus Trichomanes (Hymenophyllaceae) with special reference to stem anatomy. Bot. J. Linn. Soc. 2007, 155, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-X.; Zhang, Q.-Y.; Ebihara, A.; Iwatsuki, K. Hymenophyllaceae. In Flora of China, Pteridophytes; Wu, Z.-Y., Raven, P.H., Hong, D.-Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2013; Volumes 2–3, pp. 93–109. [Google Scholar]
- Schneider, H. Evolutionary morphology of ferns (monilophytes). Annu. Plant Rev. 2013, 45, 115–140. [Google Scholar] [CrossRef]
- Brownsey, P.J.; Perrie, L.R. Hymenophyllaceae. In Flora of New Zealand—Ferns and Lycophytes, Fascicle 16; Breitwieser, I., Heenan, P.B., Wilton, A.D., Eds.; Manaaki Whenua Press: Lincoln, New Zealand, 2016. [Google Scholar]
- Bower, F.O. The Ferns (Filicales); Cambridge University Press: London, UK, 1926; Volume II. [Google Scholar]
- Dubuisson, J.-Y. Systematic relationships within the genus Trichomanes sensu lato (Hymenophyllaceae, Filicopsida): Cladistic analysis based on anatomical and morphological data. Bot. J. Linn. Soc. 1997, 123, 265–296. [Google Scholar] [CrossRef] [Green Version]
- Pryer, K.M.; Smith, A.R.; Hunt, J.S.; Dubuisson, J.-Y. rbcL data reveal two monophyletic groups of filmy ferns (Filicopsida: Hymenophyllaceae). Am. J. Bot. 2001, 88, 1118–1130. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.; Schuettpelz, E.; Pryer, K.M.; Cranfill, R.; Magallón, S.; Lupia, R. Ferns diversified in the shadow of angiosperms. Nature 2004, 428, 553–557. [Google Scholar] [CrossRef]
- Axsmith, B.J.; Krings, M.; Taylor, T.N. A filmy fern from the Upper Triassic of North Carolina (USA). Am. J. Bot. 2001, 88, 1558–1567. [Google Scholar] [CrossRef]
- Collinson, M.E. Cainozoic ferns and their distribution. Brittonia 2001, 53, 173–235. [Google Scholar] [CrossRef]
- Taylor, T.N.; Taylor, E.L.; Krings, M. Paleobotany: The Biology and Evolution of Fossil Plants, 2nd ed.; Elsevier/Academic Press: Burlington, MA, USA; London, UK; San Diego, CA, USA; New York, NY, USA, 2009. [Google Scholar]
- Pigg, K.B.; DeVore, M.L.; Greenwood, D.R.; Sundue, M.A.; Schwartsburd, P.; Basinger, J.F. Fossil Dennstaedtiaceae and Hymenophyllaceae from the Early Eocene of the Pacific Northwest. Int. J. Plant Sci. 2021, 182, 793–807. [Google Scholar] [CrossRef]
- Deng, S.-H. Eogonocormus—A new Early Cretaceous fern of Hymenophyllaceae from China. Aust. Syst. Bot. 1997, 10, 59–67. [Google Scholar] [CrossRef]
- Vakhrameev, V.A. Stratigraphy and fossil flora of the Cretaceous deposits in Western Kazakhstan. In Regional Stratigraphy of the USSR, Part 1; Nalivkin, D.V., Ed.; Akad. Nauk SSSR: Moscow, Russia, 1952; pp. 1–340. (In Russian) [Google Scholar]
- Li, Y.; Wang, Y.-D.; Nosova, N.; Lu, N.; Xu, Y.-Y. Filmy ferns (Hymenophyllaceae) and associated spike-mosses (Selaginellaceae) from the mid-Cretaceous Kachin amber, Myanmar. Biology 2022, 11, 1629. [Google Scholar] [CrossRef]
- Fontaine, W.M. The Potomac or Younger Mesozoic Flora; US Government Printing Office: Washington, DC, USA, 1889; Volume 15, pp. 1–377.
- Knowlton, F.H. Description of a collection of Kootanie plants from the Great Falls coal field of Montana. Smithson. Misc. Collect. 1907, 50, 105–128. [Google Scholar]
- Chen, F.; Meng, X.-Y.; Ren, S.-Q.; Wu, C.-L. The Early Cretaceous Flora of Fuxin Basin and Tiefa Basin, Liaoning Province; Geological Publishing House: Beijing, China, 1988. [Google Scholar]
- Skog, J.E.; Sender, L.M. New information and family relationship (Hymenophyllaceae) for the fossil fern genus Acrostichopteris Fontaine and a new species from the Lower Cretaceous (Albian) of Spain. Am. J. Bot. 2022, 109, 1443–1455. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, D.A.; Engel, M.S.; Nascimbene, P.C. Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Am. Mus. Novit. 2002, 3361, 1–72. [Google Scholar] [CrossRef]
- Kania, I.; Wang, B.; Szwedo, J. Dicranoptycha Osten Sacken, 1860 (Diptera, Limoniidae) from the earliest Cenomanian Burmese amber. Cretac. Res. 2015, 52, 522–530. [Google Scholar] [CrossRef]
- Shi, G.-H.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.-C.; Lei, W.-Y.; Li, Q.-L.; Li, X.-H. Age constraint on Burmese amber based on U–Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Ross, A.J. Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology 2019, 2, 22–84. [Google Scholar] [CrossRef]
- Ross, A.J. Supplement to the Burmese (Myanmar) amber checklist and bibliography, 2020. Palaeoentomology 2021, 4, 57–76. [Google Scholar] [CrossRef]
- Cruickshank, R.D.; Ko, K. Geology of an amber locality in the Hukawng Valley, Northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Yu, T.-T.; Kelly, R.; Mu, L.; Ross, A.; Kennedy, J.; Broly, P.; Xia, F.-Y.; Zhang, H.-C.; Wang, B.; Dilcher, D. An ammonite trapped in Burmese amber. Proc. Natl. Acad. Sci. USA 2019, 116, 11345–11350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, J. Troubled treasure. Science 2019, 364, 722–729. [Google Scholar] [CrossRef]
- Haug, J.T.; Azar, D.; Ross, A.; Szwedo, J.; Wang, B.; Arillo, A.; Baranov, V.; Bechteler, J.; Beutel, R.; Blagoderov, V.; et al. Comment on the letter of the Society of Vertebrate Paleontology (SVP) dated April 21, 2020 regarding “Fossils from conflict zones and reproducibility of fossil-based scientific data”: Myanmar amber. Palz 2020, 94, 431–437. [Google Scholar] [CrossRef]
- Christenhusz, M.; Fay, M.; Chase, M. Plants of the World: An Illustrated Encyclopedia of Vascular Plants; Kew Publishing: Richmond, UK; Chicago University Press: Chicago, IL, USA, 2017. [Google Scholar]
- Song, Z.; Shang, Y.; Liu, Z.; Huang, P.; Wang, X.; Qian, L.; Du, B.; Zhang, D. Fossil Spores and Pollen of China, The Mesozoic Spores and Pollen; Science Press: Beijing, China, 2000; Volume 2. [Google Scholar]
- Dubuisson, J.-Y.; Péchon, T.L.; Hennequin, S.; Rouhan, G.; Salino, A.; Deblauwe, V.; Droissart, V.; Tuomisto, H.; Lehtonen, S.; Ebihara, A. New insights into the diversity, taxonomy and history of the fern genus Trichomanes (Hymenophyllaceae, Polypodiidae), with a focus on Africa and the western Indian Ocean. Bot. J. Linn. Soc. 2022, 198, 215–239. [Google Scholar] [CrossRef]
- Dubuisson, J.-Y.; Bauret, L.; Grall, A.; Li, T.; Ebihara, A.; Hennequin, S. Discussion on the taxonomy of African fern Abrodictyum rigidum (Sw.) Ebihara & Dubuisson and description of two new Abrodictyum C.Presl species (Hymenophyllaceae, Polypodiidae) for the Afro-Malagasy region. Phytotaxa 2016, 284, 151–168. [Google Scholar] [CrossRef]
- Ebihara, A.; Nitta, J.H.; Lorence, D.; Dubuisson, J.-Y. New records of Polyphlebium borbonicum, an African filmy fern, in the New World and Polynesia. Am. Fern J. 2009, 99, 200–206. [Google Scholar] [CrossRef]
- Kessler, M.; Smith, A. Prodromus of a fern flora for Bolivia. X. Hymenophyllaceae. Phytotaxa 2017, 328, 201–226. [Google Scholar] [CrossRef]
- Ponce, M.M.; Del Rio, C.; Ebihara, A.; Dubuisson, J.Y. Discussion on taxonomy of the fern genera Crepidomanes and Polyphlebium (Hymenophyllaceae) in Argentina and south-eastern South America, and description of a new local variety for Crepidomanes pyxidiferum. Bot. Lett. 2017, 164, 5–18. [Google Scholar] [CrossRef]
- Gonzatti, F.; Windisch, P.G. Synopsis of the species of the genus Polyphlebium (Hymenophyllaceae) in Brazil. Rodriguésia 2019, 70, e00532018. [Google Scholar] [CrossRef] [Green Version]
- Linnaeus, C. Species Plantarum; Impensis Laurentii Salvii: Holmiae, Sweden, 1753; Volume 2. [Google Scholar]
- Dubuisson, J.-Y.; Rouhan, G.; Grall, A.; Hennequin, S.; Senterre, B.; Pynee, K.; Ebihara, A. New insights into the systematics and evolution of the filmy fern genus Crepidomanes (Hymenophyllaceae) in the Mascarene Archipelago with a focus on dwarf species. Acta Bot. Gall. 2013, 160, 173–194. [Google Scholar] [CrossRef]
- Zotz, G. The systematic distribution of vascular epiphytes—A critical update. Bot. J. Linn. Soc. 2013, 171, 453–481. [Google Scholar] [CrossRef] [Green Version]
- Dubuisson, J.-Y.; Hennequin, S.; Rakotondrainibe, F.; Schneider, H. Ecological diversity and adaptive tendencies in the tropical fern Trichomanes L. (Hymenophyllaceae) with special reference to climbing and epiphytic habits. Bot. J. Linn. Soc. 2003, 142, 41–63. [Google Scholar] [CrossRef] [Green Version]
- Dubuisson, J.-Y.; Bary, S.; Ebihara, A.; Carnero-Diaz, E.; Boucheron-Dubuisson, E.; Hennequin, S. Epiphytism, anatomy and regressive evolution in trichomanoid filmy ferns (Hymenophyllaceae). Bot. J. Linn. Soc. 2013, 173, 573–593. [Google Scholar] [CrossRef] [Green Version]
- Hentschel, J.; Schmidt, A.R.; Heinrichs, J. Frullania cretacea sp. nov. (Porellales, Jungermanniopsida), a leafy liverwort preserved in Cretaceous amber from Myanmar. Cryptogam. Bryol. 2009, 30, 323–328. [Google Scholar]
- Heinrichs, J.; Reiner-Drehwald, M.E.; Feldberg, K.; von Konrat, M.; Hentschel, J.; Váňa, J.; Grimaldi, D.A.; Nascimbene, P.C.; Schmidt, A.R. The leafy liverwort Frullania (Jungermanniopsida) in the Cretaceous amber forest of Myanmar. Rev. Palaeobot. Palynol. 2012, 169, 21–28. [Google Scholar] [CrossRef]
- Heinrichs, J.; Schäfer-Verwimp, A.; Feldberg, K.; Schmidt, A.R. The extant liverwort Gackstroemia (Lepidolaenaceae, Porellales) in Cretaceous amber from Myanmar. Rev. Palaeobot. Palynol. 2014, 203, 48–52. [Google Scholar] [CrossRef]
- Bechteler, J.; Schmidt, A.R.; Renner, M.A.M.; Wang, B.; Pérez-Escobar, O.A.; Schäfer-Verwimp, A.; Feldberg, K.; Heinrichs, J. A Burmese amber fossil of Radula (Porellales, Jungermanniopsida) provides insights into the Cretaceous evolution of epiphytic lineages of leafy liverworts. Foss. Rec. 2017, 20, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Heinrichs, J.; Feldberg, K.; Bechteler, J.; Müller, P.; Renner, M.A.M.; Váňa, J.; Schäfer-Verwimp, A.; Schmidt, A.R. A fossil genus of the Frullaniaceae (Porellales, Jungermanniopsida) from the mid-Cretaceous of Myanmar. Cretac. Res. 2017, 74, 223–226. [Google Scholar] [CrossRef]
- Heinrichs, J.; Feldberg, K.; Müller, P.; Schäfer-Verwimp, A.; von Konrat, M.; Ilsemann, B.; Krings, M. Frullania pinnata spec. nov. (Frullaniaceae, Porellales), a new leafy liverwort in mid-Cretaceous Burmese amber from Myanmar. Cretac. Res. 2017, 78, 56–60. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.-D.; Schneider, H.; Wu, P.-C. Frullania partita sp. nov. (Frullaniaceae, Porellales), a new leafy liverwort from the mid-Cretaceous of Myanmar. Cretac. Res. 2020, 108, 104341. [Google Scholar] [CrossRef]
- Feldberg, K.; Schäfer-Verwimp, A.; Renner, M.A.M.; von Konrat, M.; Bechteler, J.; Müller, P.; Wang, Y.-D.; Schneider, H.; Schmidt, A.R. Liverworts from Cretaceous amber. Cretac. Res. 2021, 128, 104987. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.-Q.; Feldberg, K.; Wu, P.-C.; Schmidt, A.R.; Schneider, H.; Wang, Y.-D. Re-appraisal of two fossil Frullaniaceae species (Marchantiophyta, Porellales) from the mid-Cretaceous Burmese amber. Cretac. Res. 2021, 124, 104803. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.-D.; Feldberg, K.; Wang, Q.; Yang, X.-J. A new leafy liverwort of Frullania (Frullaniaceae, Porellales) from the mid-Cretaceous Kachin amber, Myanmar. Geol. J. 2021, 56, 5046–5057. [Google Scholar] [CrossRef]
- Feldberg, K.; Schäfer-Verwimp, A.; Li, Y.; Renner, M.A.M. Extending the diversity of the bryoflora in Kachin amber (Myanmar), with the description of Radula patrickmuelleri, sp. nov. and R. tanaiensis, sp. nov. (Jungermanniopsida, Porellales, Radulaceae). Foss. Rec. 2022, 25, 213–230. [Google Scholar]
- Wang, Q.; Li, Y.; Feldberg, K.; Wang, Y.-D.; Yang, X.-J. Radula heinrichsii (Radulaceae, Porellales), a leafy liverwort from the mid-Cretaceous of Myanmar. Palaeoworld 2022, 31, 679–687. [Google Scholar] [CrossRef]
- Dixon, H.N. Note on a moss in amber. J. Bot. Br. Foreign 1922, 60, 149–151. [Google Scholar]
- Bell, N.E.; York, P.V. Vetiplanaxis pyrrhobryoides, a new fossil moss genus and species from Middle Cretaceous Burmese amber. Bryologist 2007, 110, 514–520. [Google Scholar] [CrossRef]
- Hedenäs, L.; Heinrichs, J.; Schmidt, A.R. Bryophytes of the Burmese amber forest: Amending and expanding the circumscription of the Cretaceous moss genus Vetiplanaxis. Rev. Palaeobot. Palynol. 2014, 209, 1–10. [Google Scholar] [CrossRef]
- Heinrichs, J.; Schäfer-Verwimp, A.; Hedenäs, L.; Ignatov, M.S.; Schmidt, A.R. An acrocarpous moss in Cretaceous amber from Myanmar. Cretac. Res. 2014, 51, 260–265. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.-D.; Feldberg, K.; Wang, S.; Shi, C.; Cui, Y.-M.; Zhang, X.-Q. New insights into the moss genus Vetiplanaxis with a description of V. obtusus sp. nov. from the mid-Cretaceous Kachin amber, Myanmar. Rev. Palaeobot. Palynol. 2022, 301, 104659. [Google Scholar] [CrossRef]
- Poinar, G.O.; Lambert, J.B.; Wu, Y.-Y. Araucarian source of fossiliferous Burmese amber: Spectroscopic and anatomical evidence. J. Bot. Res. Inst. Texas 2007, 1, 449–455. [Google Scholar]
- Copeland, E.B. Fern evolution in Antarctica. Philipp. J. Sci. 1939, 70, 157–189. [Google Scholar]
- Iwatsuki, K. Distribution of the filmy ferns in Palaeotropics. In Tropical Botany; Larsen, K., Holm-Nielsen, L.B., Eds.; Academic Press: London, UK, 1979. [Google Scholar]
- Dubuisson, J.-Y.; Hennequin, S.; Douzery, E.J.P.; Cranfill, R.B.; Smith, A.R.; Pryer, K.M. rbcL Phylogeny of the fern genus Trichomanes (Hymenophyllaceae), with special reference to Neotropical taxa. Int. J. Plant Sci. 2003, 164, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Hennequin, S.; Ebihara, A.; Ito, M.; Iwatsuki, K.; Dubuisson, J.-Y. Molecular systematics of the fern genus Hymenophyllum s.l. (Hymenophyllaceae) based on chloroplastic coding and noncoding regions. Mol. Phylogenet. Evol. 2003, 27, 283–301. [Google Scholar] [CrossRef]
- Tenison-Woods, J.E. On the fossil flora of the coal deposits of Australia. Proc. Linn. Soc. New South Wales 1883, 8, 37–167. [Google Scholar] [CrossRef]
- Zeiller, R. Flore Fossile du Bassin Houiller de Valenciennes; Maison Quantin: Paris, France, 1886. [Google Scholar]
- Jongmans, W.; Dijkstra, S.J. Pars 43. Filicales, Pteridospermae, Cycadales. In Fossilium Catalogus. II. Plantae; Jongmans, W., Ed.; Uitgeverij Dr W. Junk: ‘s-Gravenhage, The Netherlands, 1960; pp. 1357–1452. [Google Scholar]
- Jongmans, W.; Dijkstra, S.J. Pars 63. Filicales, Pteridospermae, Cycadales. In Fossilium Catalogus. II. Plantae; Dijkstra, S.J., Ed.; Uitgeverij Dr W. Junk: ‘s-Gravenhage, The Netherlands, 1965; pp. 3421–3516. [Google Scholar]
- Menéndez, C.A. Paleophytología Kurtziana. III. 8. La flora del Cretácico Superior de Cerro Guido, Chile (1–2). Ameghiniana 1972, 9, 209–212. [Google Scholar]
- Seward, A.C. Fossil Plants, a Text-Book for Students of Botany and Geology; Cambridge University Press: Cambridge, UK, 1910; Volume II. [Google Scholar]
- Scotese, C.R. An atlas of phanerozoic paleogeographic maps: The seas come in and the seas go out. Annu. Rev. Earth. Planet. Sci. 2021, 49, 679–728. [Google Scholar] [CrossRef]
- Xi, D.-P.; Wan, X.-Q.; Li, G.-B.; Li, G. Cretaceous integrative stratigraphy and timescale of China. Sci. China Earth Sci. 2019, 62, 256–286. (In Chinese) [Google Scholar] [CrossRef]
- Dong, C.; Shi, G.; Herrera, F.; Wang, Y.; Wang, Z.; Zhang, B.; Xu, X.; Herendeen, P.S.; Crane, P.R. Leaves of Taxus with cuticle micromorphology from the Early Cretaceous of eastern Inner Mongolia, Northeast China. Rev. Palaeobot. Palynol. 2022, 298, 104588. [Google Scholar] [CrossRef]
- Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.X. The ICS international chronostratigraphic chart. Episodes 2013, 36, 199–204. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Sterile lamina fragments of Trichomanes angustum comb. nov. from mid-Cretaceous Kachin amber. PB200744. (A) Lateral view of the amber showing four lamina fragments. PB200744a–d. (B) A tripinnatifid lamina fragment showing the rachis winged only at the apex of lamina. Holotype PB200744a. (C) A lamina fragment. PB200744d. (D) A lamina fragment. PB200744h. (E) Lamina fragments. PB200744b, c. (F) Enlargement of lamina showing cell shape and arrangement. PB200744c.
Figure 1.
Sterile lamina fragments of Trichomanes angustum comb. nov. from mid-Cretaceous Kachin amber. PB200744. (A) Lateral view of the amber showing four lamina fragments. PB200744a–d. (B) A tripinnatifid lamina fragment showing the rachis winged only at the apex of lamina. Holotype PB200744a. (C) A lamina fragment. PB200744d. (D) A lamina fragment. PB200744h. (E) Lamina fragments. PB200744b, c. (F) Enlargement of lamina showing cell shape and arrangement. PB200744c.

Figure 2.
Fertile lamina fragments of Trichomanes angustum comb. nov. and its sterile lamina portions from the mid-Cretaceous Kachin amber. PB201715. (A,B) Two fertile lamina fragments in adaxial and abaxial views. PB201715a, b. (C) Apical portion of a sterile pinna. PB201715a. (D) Enlargement of lamina showing cell shape and arrangement. PB201715a.
Figure 2.
Fertile lamina fragments of Trichomanes angustum comb. nov. and its sterile lamina portions from the mid-Cretaceous Kachin amber. PB201715. (A,B) Two fertile lamina fragments in adaxial and abaxial views. PB201715a, b. (C) Apical portion of a sterile pinna. PB201715a. (D) Enlargement of lamina showing cell shape and arrangement. PB201715a.

Figure 3.
Fertile pinnae and sori of Trichomanes angustum comb. nov. from mid-Cretaceous Kachin amber. PB201715b. (A,B) A fertile pinna in abaxial and adaxial views. (C) Enlargement of pinna in adaxial view showing apical portions of sori. (D) Enlargement of pinna in abaxial view showing basal portions of sori. (E) Several sori in lateral view. (F,G) Apical portions of two sori showing non-dilated involucre mouths and filiform, long exserted receptacles.
Figure 3.
Fertile pinnae and sori of Trichomanes angustum comb. nov. from mid-Cretaceous Kachin amber. PB201715b. (A,B) A fertile pinna in abaxial and adaxial views. (C) Enlargement of pinna in adaxial view showing apical portions of sori. (D) Enlargement of pinna in abaxial view showing basal portions of sori. (E) Several sori in lateral view. (F,G) Apical portions of two sori showing non-dilated involucre mouths and filiform, long exserted receptacles.

Figure 4.
Schematic drawing of fertile pinnule of Trichomanes angustum comb. nov.

Figure 5.
Associated pollen grains adhering on lamina surface of Trichomanes angustum comb. nov. PB201715a. (A) Lamina portion showing adhering pollen grains. (B) Enlargement of lamina showing adhering pollen grains. (C–F) Individual monoporate pollen grain.
Figure 5.
Associated pollen grains adhering on lamina surface of Trichomanes angustum comb. nov. PB201715a. (A) Lamina portion showing adhering pollen grains. (B) Enlargement of lamina showing adhering pollen grains. (C–F) Individual monoporate pollen grain.

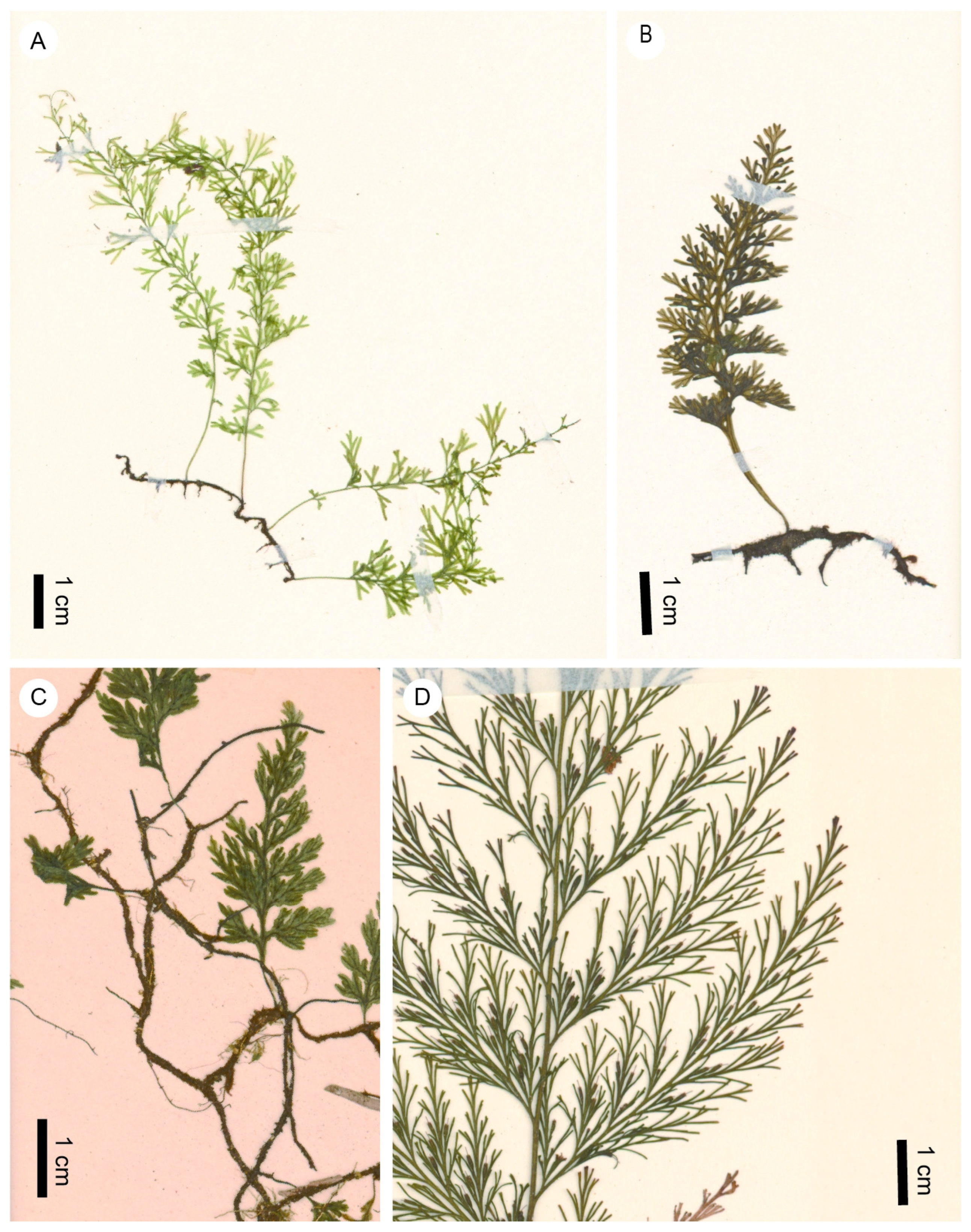
Figure 6.
Examples of extant species of Trichomanes s.l. (A) Polyphlebium angustatum (Carmich.) Ebihara et Dubuisson (TNS VS-1310151, Brazil). (B) Vandenboschia nipponica (Nakai) Ebihara (TNS VS-1222341, Japan). (C) Crepidomanes schmidtianum (Zenker ex Taschner) K.Iwats. (subg. Crepidomanes) (TNS VS-1032081, Japan). (D) Crepidomanes thysanostomum (Makino) Ebihara et K.Iwats. (subg. Nesopteris) (TNS VS-9506554, Japan).
Figure 6.
Examples of extant species of Trichomanes s.l. (A) Polyphlebium angustatum (Carmich.) Ebihara et Dubuisson (TNS VS-1310151, Brazil). (B) Vandenboschia nipponica (Nakai) Ebihara (TNS VS-1222341, Japan). (C) Crepidomanes schmidtianum (Zenker ex Taschner) K.Iwats. (subg. Crepidomanes) (TNS VS-1032081, Japan). (D) Crepidomanes thysanostomum (Makino) Ebihara et K.Iwats. (subg. Nesopteris) (TNS VS-9506554, Japan).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Y.; Ebihara, A.; Nosova, N.; Tan, Z.-Z.; Cui, Y.-M. First Fossil Record of Trichomanes sensu lato (Hymenophyllaceae) from the Mid-Cretaceous Kachin Amber, Myanmar. Life 2023, 13, 1709. https://doi.org/10.3390/life13081709
AMA Style
Li Y, Ebihara A, Nosova N, Tan Z-Z, Cui Y-M. First Fossil Record of Trichomanes sensu lato (Hymenophyllaceae) from the Mid-Cretaceous Kachin Amber, Myanmar. Life. 2023; 13(8):1709. https://doi.org/10.3390/life13081709
Chicago/Turabian StyleLi, Ya, Atsushi Ebihara, Natalya Nosova, Zhen-Zhen Tan, and Yi-Ming Cui. 2023. "First Fossil Record of Trichomanes sensu lato (Hymenophyllaceae) from the Mid-Cretaceous Kachin Amber, Myanmar" Life 13, no. 8: 1709. https://doi.org/10.3390/life13081709
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.