Plants in the Genus Tephrosia: Valuable Resources for Botanical Insecticides



1
A Key Laboratory of Natural Pesticide and Chemical Biology, Ministry of Education, South China Agricultural University, Guangzhou 510642, China
2
Department of Environmental Horticulture and Mid-Florida Research and Education Center, Institute of Food and Agricultural Sciences, University of Florida, Apopka, FL 32703, USA
*
Authors to whom correspondence should be addressed.
Insects 2020, 11(10), 721; https://doi.org/10.3390/insects11100721
Submission received: 1 October 2020
/
Revised: 16 October 2020
/
Accepted: 19 October 2020
/
Published: 21 October 2020
(This article belongs to the Special Issue Natural Substances against Insect Pests: Assets and Liabilities)
Abstract
:Simple Summary
There is an increasing interest in botanical insecticides worldwide. Plants from the genus Tephrosia are rich in bioactive phytochemicals, particularly rotenoids which include rotenone, deguelin, rotenolone, and tephrosin. Rotenoids have strong insecticidal activities against a wider range of pests. However, there has been no treatise thus far focusing on Tephrosia as insecticidal plants. This article is intended to review phytochemicals produced by selected species, their insecticidal activities, and the current status on the use of Tephrosia as botanical insecticidal plants for insect pest control.
Abstract
Synthetic insecticides are effective in controlling insect pests but can also harm nontarget organisms and the environment. During the last 40 years, there has been an increasing interest in alternative insecticides, particularly those derived from plants, commonly known as botanical insecticides. However, commercially available botanical insecticides remain limited. Rotenone is one of the earliest identified compounds and was used as fish poison and pest management. Due to its link with Parkinson disease, the use of rotenone was banned in many developed countries. Rotenone used to be isolated from Derris spp. and Lonchocarpus spp., and it can also be isolated from Tephrosia species. In this article, we present basic botanical information on selected Tephrosia species and their major compounds related to insecticidal activities and highlight the current use of extracts derived from some species, Tephrosia vogelii in particular, for control of insect pests in stored grains and crop production. The crude extracts contain multiple bioactive compounds, mainly rotenone, deguelin, rotenolone, and tephrosin, which act in either additive or synergistic fashion, resulting in effective control of insect pests. There are about 400 species in the genus Tephrosia, and species and even strains or variants vary greatly in these active compounds. We argue that a systematic evaluation of bioactive compounds in different species are needed, and species or strains with high insecticidal activities should be selected for use in the sustainable control of insect pests.
Keywords:
botanical pesticides; deguelin; insects; rotenolone; rotenone; tephrosin; Tephrosiavogelii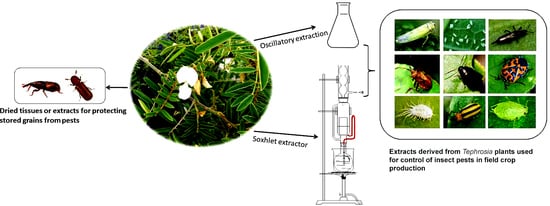
1. Introduction
Application of synthetic insecticides is the most common way of controlling insect pests in crop production [1]. The use of insecticides is credited for protecting crops from insect damage and enhancing crop productivity. It was estimated that without the use of pesticides, global food production loss would be 35–45% [2]. However, since the publication of Silent Spring [3], the application of synthetic insecticides has become one of the most controversial topics and remains the forefront regulatory issue in many countries. Insecticides are toxic, and indiscriminate use has led to the contamination of air, crop products, soil, and water; resurgence and resistance of insect pests; and toxicity to nontarget organisms [4,5,6].
An alternative to synthetic insecticides is botanical insecticides. Botanical insecticides are those consisting of dried, ground plant materials, crude plant extracts, or chemicals isolated from plants and used for managing insect pests [7]. Generally, plant secondary metabolites, like alkaloids, non-protein amino acids, steroids, phenols, flavonoids, glycosids, glucosinolates, quinones, tanins, and terpenoids are responsible for the protective action against insects. The use of botanical insecticides has a long history [7,8]. Pyrethrum was used in 400 BC. The first botanical insecticide was nicotine, dated back to the 17th century. In the mid of the 1800s, another botanical insecticide known as rotenone was introduced. Subsequently, sabadilla and other botanical insecticides were introduced [7,8]. In addition to the use of specific compounds isolated from plants, farmers across the world have used plants or plant extracts that have particularly potent defensive chemicals for controlling pests in the field and stored grains [9,10]. The modes of action of those plant extracts vary greatly including (a) confusants, phytochemicals that are unequivocal sign for insect pests to find their food source; (b) feeding deterrents, which are usually due to the activity of terpenes; (c) growth regulator activities, which can adversely affect insects’ growth and development and inhibit metamorphosis; (d) insecticidal activities resulting in the death on contact and/or by ingestion; and (e) repellents, some plants produce volatile compounds with bad odor or irritant, such as garlic and hot peppers to repel insect pests. Due to the occurrence of multiple active compounds in the extracts which may have different modes of action and could act additively or synergistically, the use of plant extracts would be more difficult to develop insecticide resistance [7,8,10] compared to the use of synthetic insecticides as they generally contain a single active compound with one mode of action [11,12]. Additionally, botanical insecticides are naturally occurring and act quickly and degraded rapidly, thus there is less pollution to the environment. Botanical insecticides have features of selectivity, low toxicity to livestock and natural enemies, with a few exception, low mammalian toxicity [7,8]. Botanical insecticides, particularly those plant-derived extracts from local farmers, are relatively cheaper in preparation and convenient in application with lower cost, due to the locally grown plants for local use in comparison to the use of synthetic ones [9,13].
Botanical insecticides have gained increasing attention as more than 20% of the research publications on insecticides are about botanical ones since the beginning of 1980 [8]. The most common botanical insecticides in the literature include pyrethrins, rotenone, sabadilla, ryania, nicotine, d-limonene, linalool, and neem [8]. However, commercially available botanical insecticides have been largely limited to pyrethroins and neem, to a lesser extent, limonene, chenopodium, capsicum oleorsin, and garlic oils [8]. Rotenone, one of the earliest identified compounds has not been widely used in insect management. Rotenone was initially isolated from ground roots or rhizomes of tropical legume Derris spp. and Lonchocarpus spp. The former produces up to 13% of rotenone, while the latter produces only about 5%. Derris spp. originated in tropical Asia, while Lonchocarpus spp. is native to western hemisphere. In addition to Derris and Lonchocarpus species, plants in the genus Tephrosia are also rich in rotenoid compounds. The content of rotenone in the pods of Tephrosia vogelii was 1.4% (dry weight) [14], and the amount of rotenoids varies with the T. vogelii variety and extractants [15] as the content of rotenoids in leaves varied from 0.65 to 4.25% [16]. T. vogelii is a fast-growing plant with large biomass. A yield of more than 14 metric tons of dried leaves and stems was harvested at Lewiston, North Carolina, the United States (U.S.), in a growing season [17]. Thus, plants in the genus Tephrosia have great potential for developing botanical insecticides. This article is intended to review general botanical characteristics of selected species, phytochemical production, and their potentials as botanical insecticides for insect pest control.
2. Common Species and Their Insecticidal Activities
The genus Tephrosia is a member of the family Leguminosae with about 400 species, and they are widely distributed in tropical, subtropical, and arid regions of the world [18,19,20]. It was reported that 50 species are native to equatorial Africa, of which 30 are found in Kenya; 70 are found in South Africa; 35 occur in India; and 30 are native of South America [21,22]. They are erect herbs or soft or woody shrubs with dense foliage grown to a height ranging from 0.5 to 4 m. They have the potential to restore soil fertility due to their ability to fix nitrogen [16,18]. Leaves are compound, leaflets are inverted lance-shaped or obovate, 7–15 cm long and 0.3–1 cm wide. Flowers are pea shape, white, purple or pinkish, 7 mm long, in a few-flowered, leaf-opposed, raceme-like clusters. Plants are self-pollinated and produce linear-long pods, 2.5–4.0 cm long and 3–4 mm wide. Seeds are ellipsoid, dark brown. Most species are diploid with a 2n chromosome number of 11. Species commonly associated with insecticidal activities include Tephrosia candida, (Roxb.) DC.; Tephrosia elata, Deflers; Tephrosia purpurea (L.) Pers.; Tephrosia villosa (L.) Pers.; Tephrosia virginiana (L.) Pers.; and T. vogelii Hook. f. (Table 1).
2.1. Tephrosia vogelii
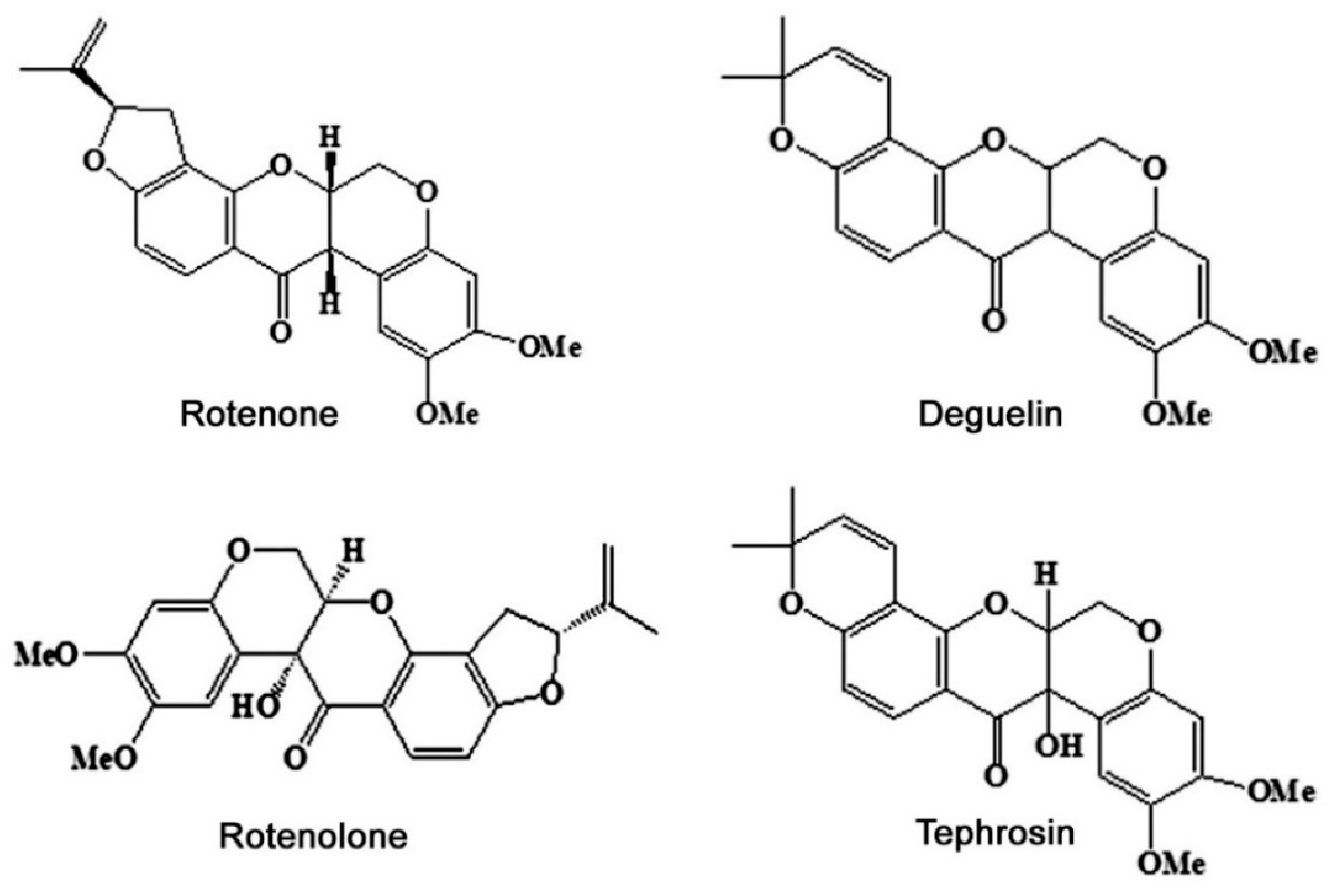
Among the 400 species, T. vogelii is the most intensively studied one. It is a herb or small tree, native to tropical Africa and can attain a height of 2 to 3 m in a growing season of 5 to 7 months. Flower color may be white, purple, or red. It produces various compounds, such as flavonoid, steroid, and rotenoids (Table 1). Rotenoids mainly include four compounds (Figure 1): (a) Rotenone, it has a molecular formula C23H22O6, crystal with a melting point of 165 °C. Aphids are killed by 3 mg/kg of rotenone in solution and oral LD50 value in rats was about 132 mg/kg body weight [23]. (b) Deguelin, a derivative of rotenone with an empirical formula of C23H22O6, the same as rotenone, crystal, and melting point is 171 °C [24]. Its LD50 value to humans ranged from 10 to 100 g [25], but such values to insects are unknown. (c) Rotenolone, crystal with a chemical formula of C23H22O7. (d) Tephrosin, a nearly colorless rotenoid with a formula of C23H22O7 and melting point of 198 °C, and it is thought to be the oxidation product of deguelin [26]. The lethality values of both rotenolone and tephrosin are unknown. The content of rotenoids in T. vogelii leaves was higher than that in petal, stems, and roots, accounting for 80% to 90% of the total rotenoids [27].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Chemical constituents in some species of Tephrosia and their insecticidal activities.
| Species | Class | Main Compound(s) | Mode of Action | Part(s) of Plant Used | Target Pests | Reference |
|---|---|---|---|---|---|---|
| Tephrosia vogelii | Rotenoid, flavonoid, and Steroid | Deguelin Tephrosin Rotenone Rotenolone α-toxicarol Elliptone Quercitin β-sitosterol Lanosterol Stigmasterol | Toxicity, larval toxicity, Antifeedant activity, Stomach or contact poison, Inhibition of oviposition, egg hatching and molting, Interference with growth and development, Interfering with the electron transport chain in mitochondria, Inhibition of cellular respiration and metabolism | Leaf Stem Root Seed Fruit coat | Acanthoscelides obtectus Aedes aegypti Aphis fabae Aphthona whitfieldi Anopheles gambiae Bactrocera curcubitea Brevicoryne brassicae Callosobruchus chinensis Callosobruchus maculatus Caryedon serratus Crocidolomia pavonana Culex quinquefasciatus Dacus cucurbitae Diabrotica undecimpunctata Euschistus heros Megalurothrips sjostedti Monolepta species Pieris rapae Phyllotreta cruciferae | [18,20,24,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48] |
| Tephrosia candida | Flavonoid and rotenoid | Candidol Dehydrorotenone 12a-hydroxyrotenone Rotenone Tephrosin Amorpholone 6a,12,-dehydodeguelin 12a-hydroxy-β-toxicarol Deguelin α-toxicarol | Stomach or contact poison, Larval toxicity, Interference with growth and development, Antifeedant activity, Inhibiting the activity of NADH oxidoreductase in electron transporters | Stem Leaf Root Seed | Aphis fabae Diaprepes abbreviates Spodoptera litura | [20,28,31,49,50,51,52,53] |
| Tephrosia elata | Flavonoid and Rotenoid | Isopongaflavone Tephrosin (S)-elatadihydrochalcone Deguelin Rotenone | Antifeedant activity, Growth inhibition activity, Inhibiting the activity of NADH, Inhibition of cellular respiration and metabolism, Larval toxicity, Inhibition of oviposition | Seed Seedpod Root | Aedes aegypti Eldana saccharina Maruca testulalis Spodoptera exempta | [20,28,54,55,56] |
| Tephrosia purpurea | Rotenoid, flavonoid, and sterol | Tephrosin Rotenone Pongaglabol Semiglabrin Quercitin Rutin (+)-tephrorins A (+)-tephrorins B β-sitosterol | Interference with growth and development, Larval toxicity, Antifeedant activity, Inhibition of cellular respiration and metabolism, Anti-inflammatory and anti-cancer properties, Inhibition of ATP production | Whole plant | Aedes aegypti Anopheles stephensi Culex quinquefasiciatus Dysdercus cingulatus Odoiporus longicollis | [20,28,57,58,59,60,61,62,63,64] |
| Tephrosia villosa | Rotenoid, sterol, and triterpenoid | Rotenone Dehydrorotenone Lupenone Stigmasterol | Larval toxicity, Stomach or contact poison, Interference with growth and development, Inhibition of oviposition Anti-parasitic | Whole plant | Anopheles gambiae Culex quinquefasciatus Tenebrio molitor Spodoptera litura | [20,47,65,66,67,68] |
| Tephrosia virginiana | Rotenoid | Rotenone Tephrosin Toxicarol | Larval toxicity, Interference with growth and development | Root | Musca domestica | [69,70] |
Rotenoids belong to advanced members of the isoflavonoid group [71]. Their biosynthesis is based on shikimic/chorismic acid pathway, covering a group of plant natural products that is based on the rotoxen skeleton [71]. Rotenone was reported to be the most toxic of the rotenoids followed by deguelin [28]. Together, the four main compounds contribute greater than 95% of the toxicity [72]. Rotenone is a contact and ingestion compound [7]. Its mode of action involves the inhibition of the electron transport at the mitochondrial level, blocking phosphorylation of ADP (adenosine di-phosphate) to ATP (adenosine triphosphate), thereby inhibiting insect metabolism [28]. It is a selective, nonsystemic insecticide with contact and stomach action and secondary acaricidal activity [73]. Rotenone has been used for centuries as a selective fish poison [28] and also as an insecticide for controlling a wide range of arthropod pests including cucumber beetle, flea beetles, harlequin bug, leafhoppers, scales, spittlebugs, squash bugs, thrips, and some fruit worms [29]. Dried leaves of T. vogelii were used to protect stored legume seeds from damage by the bruchids [30].
2.2. Tephrosia candida
Tephrosia candida, commonly known as white hoarypea, is a perennial shrub, native to India. It grows up to 3.5 m. A distinct morphological characteristic of T. candida is that it produces greater biomass than T. vogelii. T. candida produces flavonoids including candidol, candidone, ovalichalcone, dehydrorotenone, candidin, and prongachin; and rotenoids, such as tephrosin and deguelin; and sterol (Table 1) [74]. Dehydrorotenone produced in stem and leaves is a stomach or contact poison and toxic to insects [20,75]. Tephrosin isolated from roots can interfere with insect growth and development [20,49]. Deguelin occurs mainly in roots and has antifeedant and growth inhibition activities, which are used for control of insects and nematodes [20,49]. A study conducted in Malawi showed that extracts derived from T. candida for control of aphid in common bean were less effective than those extracted from T. vogelii, which was explained by the fact that active compounds in T. candida were lower than T. vogelii [31].
Tephrosia candida, however, could be used as a cover crop for repellency of larval and adult diaprepes root weevil (Coleoptera: Curculionidae). This weevil, Diaprepes abbreviates (L.), is a major threat to the sustained profitability of citrus production in Florida and the Caribbean region. Adults were deterred from feeding and produced proportionally fewer eggs when caged with foliage of T. candida compared to foliage of T. vogelii, suggesting that the leaves of T. candida might contain antifeedants with activity toward adult diaprepes root weevil [76]. Additionally, feeding damage was observed by larvae on roots of T. candida and T. vogelii, but weight gain and survival of larvae fed with T. candida were greatly reduced compared to those fed with T. vogelii. These results implied that some compounds in root of T. candida are toxic to larvae. The authors suggested that phytochemicals responsible for the antifeedant and toxic properties of T. candida toward D. abbreviates are not shared by T. vogelii, and T. candida could be used as a cover crop in citrus field for control of diaprepes root weevil [76]. In fact, T. candida has been widely used in mixed cropping regimes and as a fallow crop in tropical production systems in Vietnam [77] and India [78] where it is valued for its contribution to soil fertility and simultaneously for repelling insect pest [49,50,79].
2.3. Tephrosia elata
Tephrosia elata is a short-lived, bushy perennial shrub that occurs in Kenya and produces various phytochemicals (Table 1). Two antifeedants: tephrosin and isopongaflavone were isolated from T. elata [54]. Isopongaflavone was reported to be effective against bean pod borer (Maruca testulalis) and African sugar-cane borer (Eldana saccharina). Tephrosin exhibited high activity against African armyworm (Spodoptera exempta). Isolated rotenone was able to control of African armyworm, African sugar-cane borer, and bean pod borer [55]. Roots, seeds, and seedpods contain flavonoid, which has antifeedant activity to insects, and interfere with insect growth and development [20]. Novel flavonoids: (s)-elatadihydrochalcone, obovatin methyl ether, and praecansone A were identified from seedpods. Seeds and roots contain rotenoids including deguelin and rotenone [55,56], which together showed larvicidal activity against third instar mosquito larvae of Aedes aegypti [55,56].
2.4. Tephrosia purpurea
Tephrosia purpurea is a highly branched suberect herbaceous perennial, about 1.5 m in height with spreading branches [80]. The plant grows abundantly in the upper Gangetic plains, and western Himalayas. More than 44 phytochemicals have been structurally identified from this species, which include rotenoids, flavanols, glycosides, isoflavones, sterols, and chalcones (Table 1) [81]. T. purpurea is known for its strong insecticidal efficacy. Sahayaraj [57] evaluated the potential of T. purpurea essential oil from stem and roots. Hexadecanoic acid was found to be the most abundant compound present. Essential oil showed strong repellent activity for males compared with females of banana stem weevil (Odoiporus longicollis), a serious pest of banana. The insecticidal efficacy was attributed to the presence of compounds like rotenone and hexadecanoic acid. T. purpurea leaf extract was able to control first to fourth instars larvae and pupae of A. aegypti [82]. Furthermore, the whole plant extract of T. purpurea was tested for its larvicidal activity against the larvae of Culex quinquefasciatus. The extract showed 100% mortality in very small doses suggesting its beneficial use in controlling the mosquito reproduction [58]. Winter season is more suitable for collection of plant materials due to the presence of high content of rotenone in this season [83].
2.5. Tephrosia villosa
Tephrosia villosa is a multibranched, perennial herb, up to 90 cm high, densely clothed with white, silky hair, found in India. Roots and seedpods produce flavonoids, including tephcalostaan, villosin, and tephrinone (Table 1). Whole plants contain rotenoids, dehydrorotenone, prenylated flavonone. The ethanol extract of roots, leaves, fruit, and twigs of T. villosa showed significant activity against southern house mosquito (Culex quinquefasciatus) larvae [65]. A defensin (TvD1) isolated from T. villosa showed inhibitory activities to mealworm (Tenebrio molitor) [84]. Plant defensin, a small, cationic, cysteine-rich broad-spectrum antimicrobial peptide, has four or five disulfide bridges and has been shown to be a component of the innate immunity system in plants. Over expression of defensin gene (TvD1) in tobacco exhibited strong activity against first and second instar larvae of taro caterpillar (Spodoptera litura), an important polyphagous insect attaching 44 families of economically important plants [66].
2.6. Tephrosia virginiana
Tephrosia virginiana, commonly known as devil’s shoestring as its roots are very long and stingy, such that they can be used for twine, occurs only in North America, ranging from Texas in the southwest to Florida in the east, north to Ontario, and west to Nebraska. Roots of this species were used as piscicide by Native Americans [85]. The roots contain rotenone, tephrosin, and toxicarol (Table 1) [69]. Root extracts are toxic to aphids, houseflies, potato beetle, fleas, and lice infecting dogs and poultry. Rotenone content in roots extracts harvested at the full-bloom stage was the highest compared to those harvested at dominant, emergence, and mature seed stages, and the extracts were highly toxic to houseflies (Musca domestica L.) [70]. Detailed information on phytochemicals of this species, however, have not been reported thus far.
3. The Use of Tephrosia Plants for Managing Insect Pests
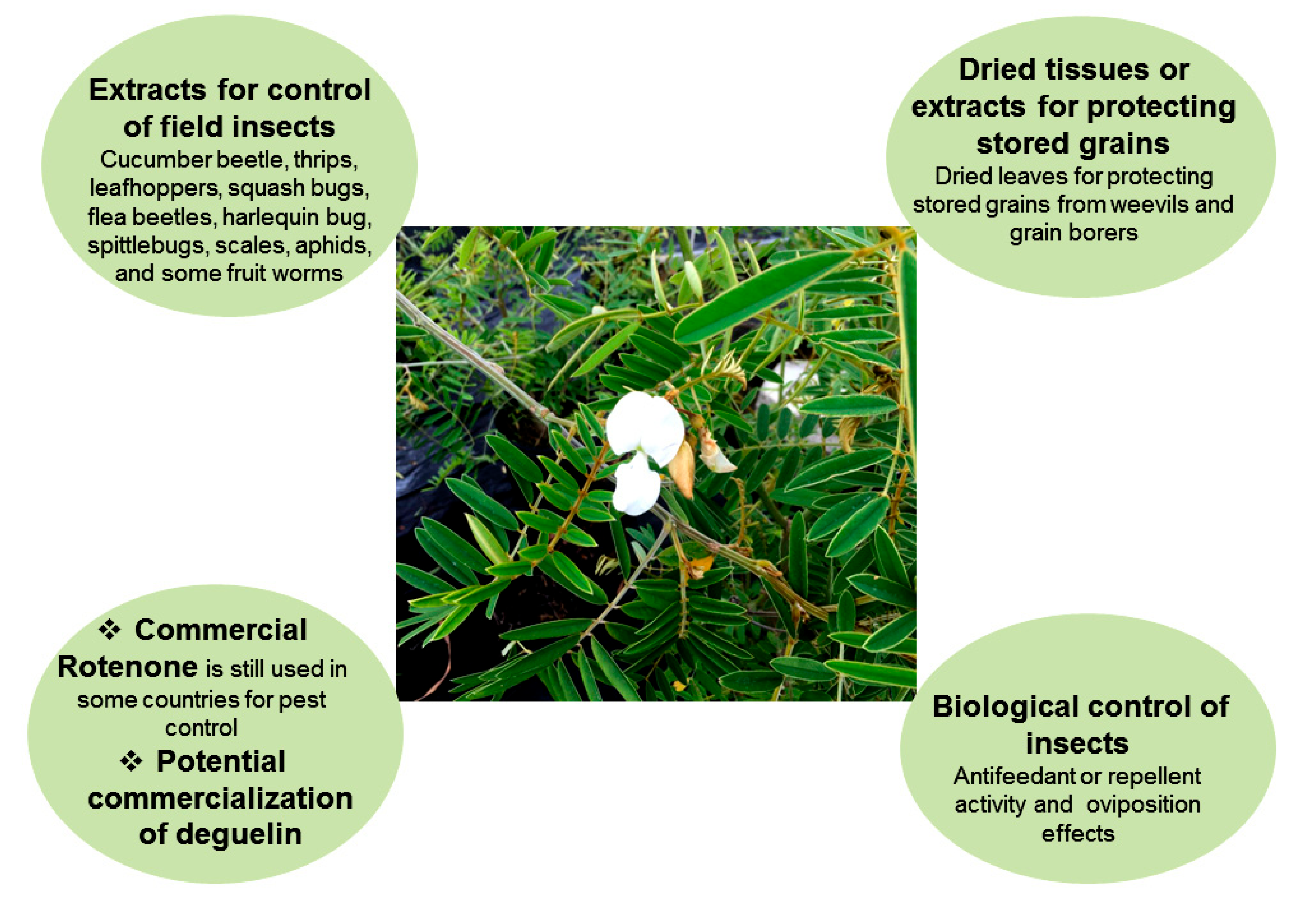
The discussed Tephrosia species are rich in bioactive compounds and show insecticidal activities against different insect pests. Based on the current information, the use of Tephrosia plants for insect pest management can be summarized as the follows (Figure 2).
3.1. Commercial Formulation of Rotenone
Commercially, rotenone is generally extracted from the roots of cube plants (Lonchocarpu utilis) and barbasco (Lonchocarpu urucu) which was referred to as Cube resin as well as from derris plants (Derris elliptica). Rotenone is also extracted from Tephrosia spp and Dalbergia paniculata [28]. Although rotenone content in cube and derris plants is higher, about 5% in dried derri roots, the cultivation of these plants is difficult because of the liana type of growth and the labor involved in harvesting the small fibrous roots. On the other hand, most Tephrosia species have large biomass. A yield of over 14 metric tons of dried leaves and stems of T. vogelii could be obtained per hectare in the U.S. [17]. Regarding the content of rotenone in plant organs, leaflets contain 80% to 90% of rotenone [17]. Additionally, breeding effort by the U.S. Department of Agriculture (USDA) in the 1970s showed that some breeding lines could have rotenone content of more than 4.5% in leaflets [17]. Thus, extraction of rotenone from T. vogelii plants could be much easier and more convenient than from cube or derri plants.
Most commercial products of rotenone come from Central and South America [28]. Rotenone almost insoluble in water, and very soluble in many organic solvents, such as ethanol, acetone, chloroform, and ether [86]. Rotenone is unstable in light and air, and not environmentally persistent [87]. It degrades rapidly under natural conditions [24]. Rotenone powders lose much of their toxicity within weeks. Thus, its storage must be protected from air, light, and alkali, and the storage temperature should not exceed 25 °C. Solutions of rotenone in organic solvent, when exposed to light and air, become successively yellow, orange, and finally deep red due to oxidation [88]. Rotenone controls aster beetles, aphids, cabbage worms, cucumber beetles, Japanese beetles, and other insects [29]. Insects poisoned by rotenone experience a drop in oxygen consumption, respiratory depression, and ataxia, which lead to convulsions, paralysis, and death by respiratory arrest [73]. Due to its sensitive to air, light, and temperature, rotenone should be applied during cloudy sky or evening with appropriate dosage to maximizing insect control efficacy and reduce application frequency [10].
Rotenone was a registered pesticide in the U.S. under the Federal Insecticide Fungicide Rodenticide Act in 1947. Its formulations include crystalline preparations (about 95%), dust (0.75%), and emulsifiable solutions (about 50%). Rotenone is also formulated with other pesticides, such as pyrethrins, carbaryl, lindane, piperonyl butoxide, and others in products to control insects, mites, ticks, lice, spiders, and undesirable fish [28]. However, due to its potentially adverse impacts on aquatic ecosystems [89] and more recently the link to Parkinson’s disease in human beings [90,91], the U.S. has banned all uses of rotenone except as a piscicide since 2012, and the European Union (EU) began a phase out of rotenone in 2008 [92]. The Codex Alimentarius Commission in 2009 proposed to remove rotenone from the list of approved substances for plant protection, which was supported by Argentina, Japan, and Kenya but opposed by Australia, Brazil, Iran, Mexico, the Philippines, Thailand, U.S., and the International Federation of Organic Agriculture Movements (IFOAM) [93]. However, rotenone is still being used in China for controlling insect pests during vegetable production, such as head cabbage with the maximum residue limit set at 0.5 mg/kg [94].
Globally, the demand for natural pesticides is growing [7,8] due to the increasing interest in organically produced safe food. Policy changes in the U.S., EU, and some other countries about the safety data and maximum residue limits for synthetic pesticides as well as rotenone may have changed the commercial scope for botanical pesticides [95]. However, some countries, such as Brazil, China, and India have led the way in policy changes that could enabled more commercialization and use of botanical pesticides [8]. Nevertheless, the use of plant extracts for insect control has been a tradition, which remains strong across the African continent and some other regions [95].
3.2. Crude Extracts for Insect Control in Field Crop Production
Water, dilute liquid soap, or organic solvent-assisted extractions of bioactive compounds from plants continue although sophisticated procedures for extraction have been advanced. Crude or unrefined plant extracts are directly used for control of insect pests. Such practices remain in low input farming, particularly in Africa and even grow elsewhere like organic farming [96]. Insecticidal activities of the extracts could be due to the action of a single compound, additive or synergistic effects of several compounds. The combined effects have been referred to as phytocomplex [97]. A study conducted in Tanzania showed that extracts derived from T. vogelii significantly controlled aphid (Aphis fabae Scopoli), foliage beetle (Ootheca mutabilis (Schonherr) and O. benigseni Weise), and flower beetle (Epicauta albovittata Gestro and Epicauta limbatipennis Pic) in the common bean production field, and the application resulted in significantly higher yield than the control treatments [98]. Similarly, studies performed in Malawi and Tanzania showed that pest abundance (aphid, bean foliage beetle, and flower beetle) was lower when synthetic pesticides were used, while application of extracts derived from T. vogelii had relatively higher number of pests but lower than control treatments. More importantly, beneficial arthropod numbers were higher in plants treated with T. vogelii extract than those treated with synthetic insecticides, suggesting that T. vogelii extract had little effect on beneficial arthropods [96]. Such effects could be attributed to lower persistence of plant extracts and different modes of action [96]. To make the insecticide, dry powdered leaves were mixed with water containing 1% liquid soap at 10% w/v ratio for 24 h. Diluted solutions containing 1–2% of the extract were sprayed in the early evening to reduce exposure to sunlight and lessen effects on beneficial insects. Such practices have largely been used in southern and eastern Africa to control field pests rather than storage pests [99]. Previous studies indicate that T. vogelii is very effective in controlling a number of hard-to-kill field insects, including cucumber beetle, leafhoppers, squash bugs, flea beetles, harlequin bug, spittlebugs, thrips, scales, and some fruit worms [99]. Anjarwalla et al. [100] also reported the efficacy of extracts derived from T. vogelii in controlling bruchids in beans and cowpeas. Chemical analysis of T. vogelii indicated the presence of the rotenoids, including deguelin, tephrosin, and rotenone (with deguelin being the most abundant) [18,39]. These results demonstrated that the use of T. vigelii extracts to control insect pests can be as effective as synthetic insecticides in terms of crop yields, while conserving the non-target arthropods. Due to the locality of plant materials and convenience in extraction and application, the crude extracts can be more easily integrated in to agro-ecologically sustainable crop production systems [96].
3.3. Tephrosia as Cover Plant for Biocontrol of Insects and Soil Nitrogen Enrichment
Another role of Tephrosia plants is their repellency of insect pest. As mentioned above, adults of diaprepes root weevil were deterred from feeding of foliage on T. candida and produced fewer eggs, and T. candida could be used as a cover crop in citrus field for control of diaprepes root weevil [76]. Additionally, acetone extracts from leaves of T. vogelii had obvious antifeedant, inhibitory effect on growth and development and ovicidal activity, as well as acute stomach action [101]. The rotenoid compounds from T. elata showed significant antifeedant activity against Maruca testulalis, Spodoptera. exempta, and Eldana sacchariana [54]. Essential oil derived from T. purpurea showed stronger repellent activity against male banana stem weevils than the female ones [57]. The potentials of these species for repellency of insect pests deserve further investigation.
Tephrosia species are also widely used as cover crops by planting with rubber, oil palm, citrus, coffee, tea, coconut, and also annual crops [102]. Some species are reported to be green manure plants in agriculture and sometimes as a windbreak, contour hedge or shade plant because of the dense foliage and good root anchorage [103]. T. noctiflora is one of the Tephrosia species first being used as a green manure [103]. T. candida proved to be one of the most satisfactory green manures because it can flourish in poor soil for several years and has a dense foliage [103]. T. candida fallows alone can raise maize grain yield by 300% [104]. In South Rwanda, T. vogelii and Cajanus cajan were intercropped with Sorghum bicolor during the long rainy season. After sorghum was harvested, green manures were cut into small pieces and incorporated together with the sorghum residues. After four years of such intercropping, results showed that only T. vogelii led to a significant increase in yields. The yield of subsequently cropped beans increased by 25% compared to the control [105].
3.4. Tephrosia for Controlling Insect Pests in Stored Grains
Tephrosia plants have long been used for protecting grains from weevils [106]. Dried leaves have the potential to protect stored legume seeds from damage by bruchids in Southern Africa [107]. The insecticidal and repellent properties of T. vogelii were tested against Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae) in stored maize grain. T. vogelii caused 85.0–93.7% insect mortality in 21 days. The mean lethal exposure times (LT50) to achieve 50% mortality varied from five to six days (7.5–10.0% w/w) to seven to eight days (2.5–5.0% w/w) [108].
Dry plant materials used for stored grains are generally recommended about 5% (v/v). However, due to the differences in the content of active compounds among varieties or even strains or variants, the application rates may vary. For example, the quantity of T. vogelii chemotype 1 (a strain or variant with higher insecticidal activity) used for control of adult bruchids could be much lower than that of chemotype 2 (a strain or variant with little insecticidal activity) [39]. The exposure of adult bruchids to chemotype 1 admixed with cowpeas led rapidly to higher levels of mortality than with chemotype 2. Interestingly, the exposure of bruchids to deguelin, the most abundant compound in the crude extract, supposedly less toxic than rotenone, caused a significantly higher mortality, while tephrosin was significantly less toxic than deguelin. The LC50 for tephrosin was calculated at approximately 200 mg/kg, whereas the LC50 values for rotenone, deguelin, sarcolobine, and toxicarol were below 10 mg/kg. The effect of obovatin 5-methyl ether against bruchids did not differ significantly from the control or the extract derived from chemotype 2 plants [39].
4. Concerns over the Use of Tephrosia Species as Botanical Insecticides
This review shows that Tephrosia plants are promising genetic resources for developing botanical insecticides. However, duo to the variable contents in active compounds among species or variants, along with incomplete understanding of their insecticidal activities, some concerns over the use of Tephrosia plants herein should be raised.
Not all species have anticipated insecticidal activities. As early as in the 1930s, Wilbaux [109] and Roark [16] suggested that only some species of Tephrosia might be the sources of rotenone. Irvine and Freyre [110] screened 16 Tephrosia species, 14 were contained some rotenoids, five contained rotenoids in their leaves, and T. vogelii had the highest leaf rotenoid content, ranging from 0.65% to 4.25%. Additionally, distinct chemotypes or variants occur in T. vogelii. Chemical analysis of plant material across Malawi identified two distinct chemotypes, one containing rotenoids for their biological activity against insects [111,112] and the other was characterized by flavones, flavanones, and flavonols [18]. Subsequent bioassays revealed that insecticidal activities of chemotype 1 were due to the presence of rotenoids, including deguelin, dehydrodeguelin, rotenone, and tephrosin; while the flavonoids in chemotype 2 were inactive [39] and had little active against insects. It was reported that about 25% of the plants of T. vogelii grown in Malawi belonged to the chemotype 2. Thus, the use of wrong species or chemotypes, such as chemotype 2 could result in ineffective in pest management as mentioned by Stevenson and Belmain [95].
Insecticidal activities may not completely rely on rotenone. In the same study [112], chemotype 1 contained deguelin as the major rotenoid along with tephrosin, and rotenone as a minor component. As mentioned above, extracts from chemotype 1 plants showed insecticidal activities, meaning deguelin plays a critical role in control of insect pests. This finding is important. Currently, it is rotenone that has been banned in several countries [92], but there has been no documentation about any regulations of deguelin. Interestingly, it has been known that plants containing high deguelin have been used as an anthelmintic agent in certain regions of China [113] and as a traditional Thai medicine to treat hepatitis and hepatic dysfunction [114]. Recently, deguelin has been shown to have potent anticancer activity against multiple cancer types and can impede carcinogenesis by enhancing cell apoptosis and preventing malignant transformation and tumor proliferation [115]. Tephrosia species or chemotypes with high deguelin contents but low levels of rotenone could be ideal plant materials for developing botanical insecticides. These results may imply that future chemical evaluation of Tephrosia species or chemotypes should focus more on deguelin. A study conducted in West Java, Indonesia showed that acetone extracts derived from leaf samples collected from seven geographical locations varied in LC50 (ranging from 0.137% to 0.371%) against cabbage head caterpillar (Crocidolomia pavonana). More importantly, compounds other than rotenone are also responsible for the insecticidal activity [116]. Early studies also showed that the leaflets of T. vogelii contained more deguelin than rotenone, the reverse was general found in the petioles, stems, and roots [17]. One variety contained deguelin but no rotenone [27]. Varietal or strain variation in active compounds should be isolated individually and labelled as different chemotypes or specific strain. They should be produced in isolation for seed production or simply propagated by vegetative means to maintain the genetic identity for further evaluation and potentially released as new cultivars for commercial production.
A major challenge in production of T. vogelii is poor seed production [17], which has affected breeding effort on developing new cultivars and large-scale production of selected desirable chemotypes or strains [17]. With the advance of plant tissue culture or microprogation [117], this problem can be easily resolved. Micropropagation through shoot culture using existing meristems can produce a large quantity of plantlets without somaclonal variation [118]. Thus, the availability of genetic identical plantlets can be used for commercial production. Due to their rapid growth characteristic, a large quantity of biomass can be used for extracting bioactive compounds for pest management.
Precautions should be taken when handling Tephrosa extracts as some species or variants contain high rotenone, and rotenone has been linked with Parkinson’s disease. Additionally, the extracts may have other potential hazardous effects to nontarget organisms even though they are much safer than commercial rotenone insecticide. The precautions include safe procedures for extraction, appropriate labeling of the extracted products, methods for storage and transportation, and safe methods for field applications at right time and correct doses.
5. Conclusions and Future Outlook
Several species of Tephrosia have long been recognized as insecticidal plants. Studies of these plants over the last 60 years have resulted in some important findings. (a) Species and varieties differ significantly in rotenoid contents. The USDA program conducted from the 1960s to 1970s clearly showed that rotenoid content varied from 0.65% to 4.25%, and hybridization and selection could further increase rotenoid content [17]. This finding suggests that rotenoid content can be improved through breeding. With the advance of molecular marker tools and omics technologies, we believe that rotenoid contents can be significantly enhanced by modern breeding technologies. (b) Chemotypes or variants occur within varieties of T. vogelii, individual plants may differ significantly in rotenone, deguelin, rotenolone, and tephrosin contents (Belmain et al., 2012). (c) Deguelin could be an important compound against insect pests. Current data indicate that deguelin could be equal to or more important than rotenone in pest management. Deguelin has been widely used as pharmaceutical compounds [115], its use as an insecticide may be environmentally more benign than rotenone. Thus, the potential of deguelin deserves future investigation. (d) Progress made on the use of T. vogelii in control of insect pests in Africa is encouraging. The use of crude extracts can control insects under the threshold level, protect other arthropods, and result in high crop yield. Such work demonstrates the effectiveness of a phytocomplex for sustainable pest control in contrast to some negative effects with the use of synthetic insecticides.
The progress made in T. vogelii calls for action to explore the rich genetic resources of Tephrosia. (a) Bioactive compounds including rotenone, deguelin, rotenolone, and tephrosin in different organs of each species should be analyzed. Based on the results, species with high levels of active compounds and insecticidal activities should be further screened for individual rotenoid contents among variants or chemotypes. (b) Chemotypes or variants with high contents of the compounds should be propagated vegetatively though in vitro shoot culture to maintain their genetic identity and fidelity. (c) These chemotypes or variants should be used as parents for hybridization to produce segregated populations for selecting progenies with even higher bioactive compounds. The developed breeding lines should be evaluated in different regions for consistence in performance, and new cultivars with high levels of a compound or a group of compounds should be released. (d) Crude extracts should be extracted from the developed cultivars, and they should be evaluated and used for control of insect pests. (e) Attention should be given to deguelin, it should be evaluated for insect control and effects on nontarget organisms and the environment. It is anticipated that deguelin could be an important compound for management of insect pests in the near future.
Author Contributions
Conceptualization, P.Z., J.C., and Z.Z.; validation, D.Q.; resource, Z.Z.; data curation, P.Z. and J.C.; writing—original draft preparation, P.Z. and J.C.; writing—review and editing, P.Z., J.C., and D.Q.; visualization, P.Z.; supervision, J.C. and Z.Z.; funding acquisition, Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Key Area Research and Development Program of Guangdong Province (No. 2020B020224002).
Acknowledgments
The authors would like to thank Terri A. Mellich for critically reviewing this manuscript.
Conflicts of Interest
This work has no conflicts of interest.
References
- Guedes, R.N.C.; Smagghe, G.; Stark, J.D.; Desneux, N. Pesticide-induced stress in arthropod pests for optimized integrated pest management programs. Annu. Rev. Entomol. 2016, 61, 43–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Carson, R. Silent Spring. Anniversary Edition; Houghton Miffin Co.: New York, NY, USA, 1962. [Google Scholar]
- Bernardes, M.F.F.; Pazin, M.; Pereira, L.C.; Dorta, D.J. Impact of pesticides on environmental and human health. In Toxicology Studies: Cells, Drugs and Environment; Andreazza, A.C., Scola, G., Eds.; InTech: Rijeka, Croatia, 2015; pp. 195–233. [Google Scholar]
- Cycon, M.; Mrozik, A.; Piotrowska-Seget, Z. Bioaugmentation as a strategy for the remediation of pesticide-polluted soil: A review. Chemosphere 2017, 172, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Campos, E.V.R.; Proença, P.L.F.; Oliveira, J.L.; Bakshi, M.; Abhilash, P.; Fraceto, L.F. Use of botanical insecticides for sustainable agriculture: Future perspectives. Ecol. Indic. 2019, 105, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Isman, M.B. Perspective botanical insecticides: For richer, for poorer. Pest Manag. Sci. 2008, 64, 8–11. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides in the twenty—first century—fulfilling the promise? Annu. Rev. Entomol. 2020, 5, 233–249. [Google Scholar] [CrossRef] [Green Version]
- Sola, P.; Mvumi, B.M.; Ogendo, J.O.; Mponda, O.; Kamanula, J.F.; Nyirenda, S.P.; Belmain, S.R.; Stevenson, P.C. Botanical pesticide production, trade and regulatory mechanisms in sub-Saharan Africa: Making a case for plant-based pesticidal products. Food Sec. 2014, 6, 369–384. [Google Scholar] [CrossRef]
- Dougoud, J.; Toepfer, S.; Bateman, M.; Jenner, W.H. Efficacy of homemade botanical insecticides based on traditional knowledge. A review. Agron. Sustain. Dev. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, N.J.; Bass, C.; Dixon, A.; Neve, P. The evolutionary origins of pesticide resistance. Biol. Rev. Camb. Philos. Soc. 2018, 94, 135–155. [Google Scholar] [CrossRef] [PubMed]
- Siegwart, M.; Graillot, B.; Lopez, C.B.; Besse, S.; Bardin, M.; Nicot, P.C.; Lopez-Ferber, M. Resistance to bioinsecticides or how to enhance their sustainability: A review. Front. Plant Sci. 2015, 6, 381. [Google Scholar] [CrossRef] [Green Version]
- Lengai, G.M.W.; Muthomi, J.W.; Mbega, E.R. Phytochemical activity and role of botanical pesticides in pest management for sustainable agricultural crop production. Sci. Afr. 2020, 7, e00239. [Google Scholar] [CrossRef]
- Hu, L.Z.; Li, X.H.; Ma, Y.Q.; Yu, X. Determination of rotenoids content in T. vogelii Hook f. Seed. J. Chin. Inst. Food Sci. Technol. 2011, 11, 206–211. [Google Scholar]
- Kariuki, D.K.; Njiru, S.N. Spectrophotometric evaluation of rotenone extraction from leaves and seeds of mature Tephrosia vogelii plant. Afr. J. Pure Appl. Chem. 2018, 12, 50–53. [Google Scholar] [CrossRef]
- Roark, R.C. Tephrosia as an Insecticide—A Review of the Literature; Roark, R.C., Ed.; U.S.Department of Agriculture, Bureau of Entomology and Plant Quarantine, Division of Insecticide Investigations, E-402: Washington, DC, USA, 1937; pp. 1–165.
- Gaskins, M.H.; White, G.A.; Martin, F.W.; Delfel, N.E.; Ruppel, E.G.; Barnes, D.K. Tephrosia vogelii: A Source of Rotenoids for Insecticidal and Piscicidal Use; Technical Bulletins from US Department of Agriculture, Economic Research Service: Washington, DC, USA, 1972. [Google Scholar] [CrossRef]
- Stevenson, P.C.; Kite, G.C.; Lewis, G.P.; Forest, F.; Nyirenda, S.P.; Belmain, S.R.; Sileshi, G.W.; Veitch, N.C. Distinct chemotypes of Tephrosia vogelii and implications for their use in pest control and soil enrichment. Phytochemistry 2012, 78, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.N.; Yan, T.; Gao, C.H.; Cao, W.H.; Huang, R.M. Natural products from the genus Tephrosia. Molecules 2014, 19, 1432–1458. [Google Scholar] [CrossRef]
- Touqeer, S.; Saeed, M.A.; Ajaib, M. A review on the phytochemistry and pharmacology of genus Tephrosia. Phytopharmacology 2013, 4, 598–637. [Google Scholar]
- Tarus, P.K.; Machocho, A.K.; Lang’at–Thoruwa, C.C.; Chhabra, S.C. Flavonoids from Tephrosia aequilata. Phytochemistry 2002, 60, 375–379. [Google Scholar] [CrossRef]
- Beentje, H. Kenya Trees, Shrubs and Lianas; National Museums of Kenya: Nairobi, Kenya, 1994; pp. 269–320. [Google Scholar]
- Morgan, E.; Wilson, I.D. Insect hormones and insect chemical ecology. In Comprehensive Natural Products Chemistry, Vol. 8: Miscellaneous Natural Products Including Marine Natural Products, Pheromones, Plant Hormones, and Aspects of Ecology; Mori, K., Ed.; Pergamon, Elsevier: Oxford, UK, 1999; pp. 263–376. [Google Scholar]
- Dzenda, T.; Ayo, J.A.; Adelaiye, A.B.; Adaudi, A.O. Ethno-medical and veterinary uses of Tephrosia vogelii Hook. F.: A review. Nig. Vet. J. 2007, 28, 24–30. [Google Scholar] [CrossRef]
- Woo, J.K.; Choi, D.S.; Tran, H.T.; Gilbert, B.E.; Hong, W.K.; Lee, H.Y. Liposomal encapsulation of deguelin: Evidence for enhanced antitumor activity in tobacco carcinogen–induced and oncogenic K-ras–induced lung tumorigenesis. Cancer Prev. Res. 2009, 2, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Murillo, G.; Salti, G.I.; Kosmeder, J.W.; Pezzuto, J.M.; Mehta, R.G. Deguelin inhibits the growth of colon cancer cells through the induction of apoptosis and cell cycle arrest. Eur. J. Cancer 2002, 38, 2446–2454. [Google Scholar] [CrossRef]
- Delfel, N.E.; Tallent, W.H.; Carlson, D.G.; Wolff, I.A. Distribution of rotenone and deguelin in Tephrosia vogelii and separation of rotenoid-rich fractions. J. Agric. Food Chem. 1970, 18, 385–390. [Google Scholar] [CrossRef]
- Ling, N. Rotenone—A review of its toxicity and use for fisheries management. Sci. Conserv. 2002, 211, 1–40. [Google Scholar]
- Kerebba, N.; Byamukama, R.; Oyedeji, A.O.; Oyedeji, O.O.; Kuria, S.K. Pesticidal activity of Tithonia diversifolia (Hemsl.) A. Gray and Tephrosia vogelii (Hook f.); phytochemical isolation and characterization: A review. S. Afr. J. Bot. 2019, 121, 366–376. [Google Scholar] [CrossRef]
- Koona, P.; Dorn, S. Extracts from Tephrosia vogelii for the protection of stored legume seeds against damage by three bruchid species. Ann. Appl. Biol. 2005, 147, 43–48. [Google Scholar] [CrossRef]
- Kayange, C.D.M.; Njera, D.; Nyirenda, S.P.; Mwamlima, L. Effectiveness of Tephrosia vogelii and Tephrosia candida extracts against common bean aphid (Aphis fabae ) in Malawi. Adv. Agric. 2019, 2019, 6704834. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Khanna, P. Production of rotenoids from Tephrosia spp. in vivo and in vitro tissue cultures. Indian J. Exp. Biol. 1975, 13, 84–85. [Google Scholar]
- Lambert, N.; Trouslot, M.F.; Nef-Campa, C.; Chrestin, H. Production of rotenoids by heterotrophic and photomixotrophic cell cultures of Tephrosia vogelii. Phytochemistry 1993, 34, 1515–1520. [Google Scholar] [CrossRef]
- Marston, A.; Msonthi, J.D.; Hostetmann, K. On the reported molluscicidal activity from Tephrosia vogelii leaves. Phytochemistry 1984, 23, 1824–1825. [Google Scholar] [CrossRef]
- Mikami, A.Y.; Ventura, M.U.; Andrei, C.C. Brown stink bug mortality by seed extracts of Tephrosia Vogelii containing deguelin and tephrosin. Braz. Arch. Biol. Technol. 2018, 61, e18180028. [Google Scholar] [CrossRef]
- Fang, N.; Casida, J.E. Anticancer action of cubé insecticide: Correlation for rotenoid constituents between inhibition of NADH: Ubiquinone oxidoreductase and induced ornithine decarboxylase activities. Proc. Natl. Acad. Sci. USA 1998, 95, 3380–3384. [Google Scholar] [CrossRef] [Green Version]
- Preston, S.; Korhonen, P.K.; Mouchiroud, L.; Cornaglia, M.; McGee, S.L.; Young, N.D.; Davis, R.A.; Crawford, S.; Nowell, C.; Ansell, B.R.E.; et al. Deguelin exerts potent nematocidal activity via the mitochondrial respiratory chain. FASEB J. 2017, 31, 4515–4532. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, J.P.; Basu, P.K. Sterols and rotenoids from fruit coat of Tephrosia vogelii Hook. Acta Hortic. 1988, 188A, 155–157. [Google Scholar]
- Belmain, S.R.; Amoah, B.A.; Nyirenda, S.P.; Kamanula, J.F.; Stevenson, P.C. Highly variable insect control efficacy of Tephrosia vogelii Chemotypes. J. Agric. Food Chem. 2012, 60, 10055–10063. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.K.; Freyre, R.H. Recovery of natural insecticides from Tephrosia vogelii. III. An improved procedure for sampling and assaying rotenoid content in leaves. Econ. Bot. 1967, 21, 93–98. [Google Scholar] [CrossRef]
- Fukami, J.I. Insecticides as inhibitors of respiration. In Approaches to New Leads for Insecticides; Part of The Proceedings in Life Sciences Book Series; von Keyserlingk, H.C., Jäger, A., von Szczepanski, C., Eds.; Springer: Berlin, Germany, 1976; pp. 47–69. [Google Scholar]
- Hu, L.Z.; Li, X.H.; Ma, Y.Q.; Yu, X. Study on bacteriostatic activity of extracts in different solvents from Tephrosia vogelii Hook f. seeds. Sci. Technol. Food Ind. 2011, 32, 85–88. [Google Scholar]
- Roark, R.C. A Review of the Insecticidal Uses of Rotenone and Rotenoids from Derris, Lonchocarpus (Cube and Timbo), Tephrosia, and Related Plants; United States Department of Agriculture, Bureau of Entomology and Plant Quarantine: Washington, DC, USA, 1944; pp. 2–31. [Google Scholar]
- Zeng, X.N.; Zhang, S.X.; Fang, J.F.; Han, J.Y. Comparison of the bioactivity of elliptone and rotenone against several agricultural insect pests. Kun Chong Xue Bao Acta Entomol. Sin. 2001, 45, 611–616. [Google Scholar]
- Kamal, R. Sterols from tissue cultures of Tephrosia species. Bangladesh Pharm. J. 1978, 7, 12–16. [Google Scholar]
- Samuel, V.J.; Mahesh, A.R.; Murugan, V. Phytochemical and pharmacological aspects of Tephrosia genus:A brief review. J. Appl. Pharm. Sci. 2019, 9, 117–125. [Google Scholar]
- Kidukuli, A.W.; Maregesi, S.M.; Saria, J.; Otieno, N.J.; Lawi, Y.; Nondo, R.S.; Innocent, E.M.; Mlimbila, J.; Mihale, M.J.; Moshi, M.J. Larvicidal efficacy of some Tephrosia species extracts against Anopheles Gambiae Ss and Culex Quinquefasciatus Say. Spatula DD 2015, 5, 21–25. [Google Scholar] [CrossRef]
- Alao, F.O.; Adebayo, T.A. Comparative efficacy of Tephrosia vogelii and Moringa oleifera against insect pests of watermelon (Citrullus lanatus Thumb). Int. Lett. Nat. Sci. 2015, 35, 71–78. [Google Scholar]
- Andrei, C.C.; Vieira, P.C.; Fernandes, J.B.; Da Silva MFGF. Dimethylchromene rotenoids from Tephrosia candida. Phytochemistry 1997, 46, 1081–1085. [Google Scholar] [CrossRef]
- Kole, R.K.; Satpathi, C.; Chowdhury, A.; Ghosh, M.R.; Adityachaudhury, N. Isolation of amorpholone, a potent rotenoid insecticide from Tephrosia candida. J. Agric. Food Chem. 1992, 40, 1208–1210. [Google Scholar] [CrossRef]
- Dutt, S.K.; Chibber, S.S. Candidol, a flavonol from Tephrosia candida. Phytochemistry 1983, 22, 325–326. [Google Scholar] [CrossRef]
- Roy, M.; Mitra, S.R.; Bhattacharyya, A.; Adityachaudhury, N. Candidone, a flavanone from Tephrosia candida. Phytochemistry 1986, 25, 961–962. [Google Scholar] [CrossRef]
- Parmar, V.S.; Jain, R.; Gupta, S.R.; Boll, P.M.; Mikkelsen, J.M. Phytochemical Investigation of Tephrosia candida: Hplc separation of tephrosin and 12a-hydroxyrotenone. J. Nat. Prod. 1988, 51, 185. [Google Scholar] [CrossRef]
- Bentley, M.D.; Hassanali, A.; Lwande, W.; Njoroge, P.E.W.; Sitayo, E.N.; Yatagai, M. Insect antifeedants from Tephrosia elata Deflers. Int. J. Trop. Insect Sci. 1987, 8, 85–88. [Google Scholar] [CrossRef] [Green Version]
- Muiva, L.M.; Yenesew, A.; Derese, S.; Heydenreich, M.; Peter, M.G.; Akala, H.M.; Eyase, F.; Waters, N.C.; Mutai, C.; Keriko, J.M.; et al. Antiplasmodial β-hydroxydihydrochalcone from seedpods of Tephrosia elata. Phytochem. Lett. 2009, 2, 99–102. [Google Scholar] [CrossRef]
- Mutisya, L.M. Antiplasmodial and Larvicidal Flavonoids from the Seedpods of Tephrosia Elata and Tephrosia Aequilata; Jomo Kenyatta University of Agriculture and Technology: Juja, Kenya, 2009. [Google Scholar]
- Sahayaraj, K.; Kombiah, P.; Dikshit, A.K.; Rathi, J.M. Chemical constituents of the essential oils of Tephrosia purpurea and Ipomoea carnea and their repellent activity against Odoiporus longicollis. J. Serbian Chem. Soc. 2015, 80, 465–473. [Google Scholar] [CrossRef]
- Kumar, D.; Dhamodaran, P.; Nilani, P.; Balakrishnan, N. Larvicidal activity of Tephrosia purpurea, (L) against the Larvae of Culex quinquefasiciatus. J. Appl. Pharm. Sci. 2012, 2, 219–221. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.S.; Yadav, S.S.; Singh, P.; Nandal, A.; Singh, N.; Ganaie, S.A.; Yadav, N.; Kumar, R.; Bhandoria, M.S.; Bansal, P. A comprehensive review on ethnomedicine, phytochemistry, pharmacology, and toxicity of Tephrosia purpurea (L.) Pers. Phytother. Res. 2020, 34, 1902–1925. [Google Scholar] [CrossRef]
- Ahmad, V.U.; Ali, Z.; Hussaini, S.R.; Iqbal, F.; Zahid, M.; Abbas, M.; Saba, N. Flavonoids of Tephrosia purpurea. Fitoterapia 1999, 70, 443–445. [Google Scholar] [CrossRef]
- Jain, A.; Lodhi, S.; Singhai, A.K. Simultaneous estimation of quercetin and rutin in Tephrosia purpurea Pers by high performance thin-layer chromatography. Asian J. Tradit. Med. 2009, 4, 104–109. [Google Scholar]
- Chang, L.C.; Gerhäuser, C.; Song, L.; Farnsworth, N.R.; Pezzuto, J.M.; Kinghorn, A.D. Activityguided isolation of constituents of Tephrosia purpurea with the potential to induce the phase II enzyme, quinone reductase. J. Nat. Prod. 1997, 60, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Parmar, V.S.; Rathore, J.S.; Jain, R.; Henderson, D.A.; Malone, J.F. Occurrence of pongamol as the enol structure in Tephrosia purpurea. Phytochemistry 1989, 28, 591–593. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Shoba, J. Toxic effect of Tephrosia purpurea (Linn.) and Acalypha indica (Linn.) aqueous extracts impact on the mortality, macromolecules, intestinal electrolytes and detoxication enzymes of Dysdercus cingulatus (Fab.). Asian J. Biochem. 2012, 7, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Nondo, R.S.; Mbwambo, Z.H.; Kidukuli, A.W.; Innocent, E.M.; Mihale, M.J.; Erasto, P.; Moshi, M.J. Larvicidal, antimicrobial and brine shrimp activities of extracts from Cissampelos mucronata and Tephrosia villosa from coast region, Tanzania. BMC Complement. Altern. Med. 2011, 11, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Vijayan, S.; Singh, N.K.; Shukla, P.; Kirti, P.B. Defensin (TvD1) from Tephrosia villosa exhibited strong anti-insect and antifungal activities in transgenic tobacco plants. J. Pest Sci. 2013, 86, 337–344. [Google Scholar] [CrossRef]
- Prashant, A.; Krupadanam, G.L.D. Dehydro-6-hydroxyrotenoid and lupenone from Tephrosia villosa. Phytochemistry 1993, 32, 484–486. [Google Scholar] [CrossRef]
- Ganapaty, S.; Nyamathulla, S.; Srilakshmi, G.V.K.; Prasad, R. Chemical and antimicrobial studies of the roots of Tephrosia villosa (L) Pers. Asian J. Chem. 2008, 20, 4498–4502. [Google Scholar]
- Little, V.A. Devil’s shoestring as an insecticide. Science 1931, 73, 315–316. [Google Scholar] [CrossRef]
- Sievers, A.F.; Lowman, M.S.; Russell, G.A.; Sullivan, W.N. Changes in the insecticidal value of the roots of cultivated devil’s shoestring, Tephrosia virginiana, at four seasonal growth periods. Amer. J. Bot. 1940, 27, 284–289. [Google Scholar] [CrossRef]
- Crombie, L.; Whiting, D.A. Biosynthesis of the rotenoid group of natural products: Applications of isotope methodology. Phytochemistry 1998, 49, 1479–1507. [Google Scholar] [CrossRef]
- Fang, N.; Casida, J.E. Cube resin insecticide: Identification and biological activity of 29 rotenoid constituents. J. Agric. Food Chem. 1999, 47, 2130–2136. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.C. Rotenone. In Encyclopedia of Toxicology, 3rd ed.; Philip, W., Ed.; Academic Press; Elsevier: London, UK, 2014; pp. 185–187. [Google Scholar]
- Hegazy, M.E.F.; Mohamed, A.E.H.; El-Halawany, A.M.; Djemgou, P.C.; Shahat, A.A.; Pare, P.W. Estrogenic activity of chemical constituents from Tephrosia candida. J. Nat. Prod. 2011, 74, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, F. Toxicology of Insecticides; Plenum Press; Springer: New York, NY, USA, 1975; pp. 105–251. [Google Scholar]
- Lapointe, S.L.; McKenzie, C.L.; Hunter, W.B. Toxicity and repellency of Tephrosia candida to larva and adult Diaprepes root weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 2003, 96, 811–816. [Google Scholar] [CrossRef]
- Fagerström, M.H.H.; Noordwijk, M.V.; Phien, T.; Vinh, N.C. Innovations within upland rice-based systems in northern Vietnam with Tephrosia candida as fallow species, hedgerow, or mulch: Net returns and farmers’ response. Agric. Ecosyst. Environ. 2001, 86, 23–37. [Google Scholar] [CrossRef]
- Basu, P.K.; Gupta, I. Role of Tephrosia candida DC in enhancing the amino acid content of north Bengal soil. Geobios 1988, 15, 18–21. [Google Scholar]
- Andrei, C.C.; Vieira, P.C.; Fernandes, J.B.; Silva, M.; Fo, E.R. New spirorotenoids from Tephrosia candida. Z. Naturforsch. C 2002, 57, 418–422. [Google Scholar] [CrossRef] [Green Version]
- Dalwadi, P.P.; Patel, J.L.; Patani, P.V. Tephrosia purpurea Linn (Sharpunkha, wild indigo): A review on phytochemistry and pharmacological studies. Indian J. Pharm. Biol. Res. 2014, 2, 108. [Google Scholar] [CrossRef]
- Pelter, A.; Ward, R.S.; Rao, E.V.; Raju, N.R. 8-Substituted flavonoids and 3’-substituted 7-oxygenated chalcones from Tephrosia purpurea. J. Chem. Soc. Perkin Trans. 1981, 1, 2491–2498. [Google Scholar] [CrossRef]
- Venkadachalam, R.; Subramaniyan, V.; Palani, M.; Subramaniyan, M.; Srinivasan, P.; Raji, M. Mosquito larvicidal and pupicidal activity of Tephrosia purpurea Linn. (Family: Fabaceae) and Bacillus sphaericus against, dengue vector, Aedes aegypti. Pharmacogn. J. 2017, 9, 737–742. [Google Scholar] [CrossRef] [Green Version]
- Khatoon, S.; Irshad, S.; Pandey, M.M.; Rastogi, S.; Rawat, A.K.S. A validated HPTLC densitometric method for determination of lupeol, β-sitosterol and rotenone in Tephrosia purpurea: A seasonal study. J. Chromatogr. Sci. 2019, 57, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, S.; Imani, J.; Tanneeru, K.; Guruprasad, L.; Kogel, K.H.; Kirti, P.B. Enhanced antifungal and insect a-amylase inhibitory activities of Alpha-TvD1, a peptide variant of Tephrosia villosa defensin (TvD1) generated through in vitro mutagenesis. Peptides 2012, 33, 220–229. [Google Scholar] [CrossRef]
- Gard, M. The toxicity of extracts of Tephrosia virginiana (Fabaceae) in Oklahoma. Okla. Native Plant Rec. 2010, 10, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Othman, Z.S.; Hassan, N.H.; Zubairi, S.I. Imidazolium-Based Ionic Liquid Binary Solvent System as an Extraction Medium in Enhancing the Rotenone Yield Extracted from Derris elliptica Roots. In Progress and Developments in Ionic Liquids; IntechOpen: London, UK, 2017; pp. 495–515. [Google Scholar]
- Casacchia, T.; Sofo, A.; Toscano, P.; Sebastianelli, L.; Perri, E. Persistence and effects of rotenone on oil quality in two Italian olive cultivars. Food Chem. Toxicol. 2009, 47, 214–219. [Google Scholar] [CrossRef]
- Cheng, H.M.; Yamamoto, I.; Casida, J.E. Rotenone photodecomposition. J. Agric. Food Chem. 1972, 20, 850–856. [Google Scholar] [CrossRef]
- Mangum, F.A.; Madrigal, J.L. Rotenone effect on aquatic macroinvertebrates of the Strawberry River, Utah: A five-year summary. J. Freshw. Ecol. 1999, 14, 125–135. [Google Scholar] [CrossRef]
- Sherer, T.B.; Betarbet, R.; Testa, C.M.; Seo, B.B.; Richardson, J.R.; Kim, J.H.; Miller, G.W.; Yagi, T.; Matsuno-Yagi, A.; Greenamyre, J.T. Mechanism of toxicity in rotenone models of Parkinson’s disease. J. Neurosci. 2003, 23, 10756–10764. [Google Scholar] [CrossRef]
- Tanner, C.M.; Kamel, F.; Ross, G.W.; Hoppin, J.A.; Goldman, S.M.; Korell, M.; Marras, C.; Bhudhikanok, G.S.; Kasten, M.; Chade, A.R.; et al. Rotenone, Paraquat, and Parkinson’s Disease. Environ. Health Perspect. 2011, 119, 866–872. [Google Scholar] [CrossRef] [Green Version]
- Baker, B. Rotenone Use in Organic Farming. Hygeia Anal. 2017. Available online: https://hygeia-analytics.com/2017/01/04/rotenone-use-in-organic-farming/ (accessed on 20 September 2020).
- Codex Alimentarius Commission (CAC). Guidelines for the Production, Processing, Labelling and Marketing of Organically Produced Foods: Annex 2: Deletion of Rotenone (CL2008/27-fl): Government Comments at Step 3. 2009. Available online: http://www.fao.org/input/download/standards/360/cxg_032e.pdf (accessed on 10 October 2020).
- United States Department of Agriculture (USDA). Foreign Agricultural Service. China Release Newmaximum Residue Limits for Pesticides in Food. Global Agricultural Information Network. 2018. Available online: https://apps.fas.usda.gov/newgainapi/api/report/downloadreportbyfilename?filename=China%20Releases%20Standard%20for%20Maximum%20Residue%20Limits%20in%20Foods%20_Beijing_China%20-%20Peoples%20Republic%20of_5-22-2019.pdf (accessed on 18 September 2020).
- Stevenson, P.C.; Belmain, S.R. Pesticidal plants in African agriculture: Local uses and global perspectives. Outlooks Pest Manag. 2016, 27, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Tembo, Y.; Mkindi, A.G.; Mkenda, P.A.; Mpumi, N.; Mwanauta, R.; Stevenson, P.C.; Ndakidemi, P.A.; Belmain, S.R. Pesticidal plant extracts improve yield and reduce insect pests on legume crops without harming beneficial arthropods. Front. Plant Sci. 2018, 9, 1425. [Google Scholar] [CrossRef] [PubMed]
- Donno, D.; Beccaro, G.L.; Cerutti, A.K.; Mellano, M.G.; Bounous, G. Bud extracts as new phytochemical source for herbal preparations: Quality control and standardization by analytical fingerprint. In Phytochemicals—Isolation, Characterisation and Role in Human Health; Rao, V., Rao, L., Eds.; Tech: Rijeka, Croazia, 2015; pp. 187–218. [Google Scholar]
- Mkenda, P.; Mwanauta, R.; Stevenson, P.C.; Ndakidemi, P.; Mtei, K.; Belmain, S.R. Extracts from field margin weeds provide economically viable and environmentally benign pest control compared to synthetic pesticides. PLoS ONE 2015, 10, e0143530. [Google Scholar] [CrossRef] [PubMed]
- Reuben, S.O.; Masunga, M.; Makundi, R.; Misangu, R.N.; Kilonzo, B.; Mwatawala, M.; Lyimo, H.F.; Ishengoma, C.G.; Msuya, D.G.; Mulungu, L.S. Control of Cowpea Weevil (Callosobruchus maculatus L.) in Stored Cowpea (Vigna unguiculatus L.) Grains using Botanicals. Asian J. Plant Sci. 2006, 5, 91–97. [Google Scholar]
- Anjarwalla, P.; Ofori, D.A.; Jamnadass, R.; Mowo, J.G.; Stevenson, P.C. Proceedings of the Training Workshop on Sustainable Production, Harvesting and Conservation of Botanical Pesticides; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2013; Available online: http://projects.nri.org/options/images/handbook.pdf (accessed on 18 September 2020).
- Huang, J.G.; Zhou, L.J.; Xu, H.H.; An, Y.X. Recent advances in the insecticidal plant, Tephrosia vogelii. Plant Prot. 2006, 32, 18–21. [Google Scholar]
- Baligar, V.C.; Fageria, N.K. Agronomy and physiology of tropical cover crops. J. Plant Nutr. 2007, 30, 1287–1339. [Google Scholar] [CrossRef]
- Bosman, M.T.M.; DeHaas, A.J.P. A revision of the genus Tephrosia (Leguminosae-Papilionoideae) in Malesia. Blumea 1983, 28, 421–487. [Google Scholar]
- Munthali, M.G.; Gachene, C.K.K.; Sileshi, G.W.; Karanja, N.K. Amendment of Tephrosia improved fallows with inorganic fertilizers improves soil chemical properties, N uptake, and maize yield in Malawi. Int. J. Agron. 2014. [Google Scholar] [CrossRef] [Green Version]
- Hagedorn, F.; Steiner, K.G.; Sekayange, L.; Zech, W. Effect of rainfall pattern on nitrogen mineralization and leaching in a green manure experiment in South Rwanda. Plant Soil 1997, 195, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, P.C.; Arnold, S.E.J.; Belmain, S.R. Pesticidal plants for stored product pests on small-holder farms in Africa. In Advances in Plant Biopesticides; Springer: Berlin/Heidelberg, Germany, 2014; pp. 149–172. [Google Scholar] [CrossRef]
- Boeke, S.J.; van Loon, J.J.A.; van Huis, A.; Kossou, D.K.; Dicke, M. The Use of Plant Material to Protect Stored Seeds against Seed Beetles: A Review; Laboratory of Entomology, Wageningen University: Wageningen, The Netherlands, 2001; Available online: https://edepot.wur.nl/282994 (accessed on 20 September 2020).
- Ogendo, J.O.; Belmain, S.R.; Deng, A.L.; Walker, D.J. Comparison of toxic and repellent effects of Lantana Camara L. with Tephrosia vogelii hook and a synthetic pesticide against Sitophilus zeamais motschulsky (Coleoptera: Curculionidae) in stored maize grain. Int. J. Trop. Insect Sci. 2003, 23, 127–135. [Google Scholar] [CrossRef]
- Wilbaux, R. Considerations sur Tephrasia vogelii Hook. f. et uncertain hombre d’especes voisines. Ann. Gembloux. 1935, 41, 1–30. [Google Scholar]
- Irvine, J.E.; Freyre, R.H. The occurrence of rotenoids in some species of the genus Tephrosia. J. Agric. Food Chem. 1959, 7, 106–107. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Mkindi, A.G.; Tembo, Y.; Mbega, E.R.; Medvecky, B.; Kendal-Smith, A.; Farrell, I.W.; Ndakidemi, P.A.; Belmain, S.R.; Stevenson, P.C. Phytochemical analysis of Tephrosia vogelii across East Africa reveals three chemotypes that influence its use as a pesticidal plant. Plants 2019, 8, 597. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.Y.; Chen, L.J.; Li, Y.F.; Peng, A.H.; Fu, A.F.; Song, H.; Tang, M.H.; Luo, H.D.; Luo, Y.F.; Xu, Y.B.; et al. Preparative isolation and purification of three rotenoids and one isoflavone from the seeds of Millettia pachycarpa Benth by high-speed counter-current chromatography. J. Chromatogr. A 2008, 1178, 101–107. [Google Scholar] [CrossRef]
- Wen, L.; Chen, Y. Molecular mechanism of deguelin in anti-tumor effect. Curr. Pharm. Anal. 2012, 8, 14–19. [Google Scholar] [CrossRef]
- Varughese, R.S.; Lam, W.S.-T.; Marican, A.A.B.H.; Viganeshwari, S.H.; Bhave, A.S.; Syn, N.L.; Wang, J.G.; Wong, A.L.-A.; Kumar, A.P.; Lobie, P.E.; et al. Biopharmacological considerations for accelerating drug development of deguelin, a rotenoid with potent chemotherapeutic and chemopreventive potential. Cancer 2019, 125, 1789–1798. [Google Scholar] [CrossRef]
- Prijono, D.; Panggraito, A.; Syahbirin, G. Variation in insecticidal activity of leaf extracts of Tephrosia vogelii J.D. Hooker (Leguminosae) from West Java, Indonesia. In Proceedings of the 3rd International Conference on Applied Life Sciences (ICALS2014), Bangi, Malaysia, 18–20 September 2014; ISALS Publishing: London, UK, 2014; pp. 1–5. [Google Scholar]
- Chen, J.; Henny, R.J. Role of micropropagation in the development of ornamental foliage plant industry. In Floriculture, Ornamental and Plant Biotechnology, V; Da Silva, J.A.T., Ed.; Global Science Books: London, UK, 2008; pp. 206–218. [Google Scholar]
- Chen, J.; Henny, R.J. Somaclonal variation: An important source for cultivar development of floriculture crops. In Floriculture, Ornamental and Plant Biotechnology, II; Da Silva, J.A.T., Ed.; Global Science Books: London, UK, 2006; pp. 244–253. [Google Scholar]
Figure 1.
Major chemical constituents in Tephrosia vogelii plants.

Figure 2.
The use of Tephrosia plants for pest management.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, P.; Qin, D.; Chen, J.; Zhang, Z. Plants in the Genus Tephrosia: Valuable Resources for Botanical Insecticides. Insects 2020, 11, 721. https://doi.org/10.3390/insects11100721
AMA Style
Zhang P, Qin D, Chen J, Zhang Z. Plants in the Genus Tephrosia: Valuable Resources for Botanical Insecticides. Insects. 2020; 11(10):721. https://doi.org/10.3390/insects11100721
Chicago/Turabian StyleZhang, Peiwen, Deqiang Qin, Jianjun Chen, and Zhixiang Zhang. 2020. "Plants in the Genus Tephrosia: Valuable Resources for Botanical Insecticides" Insects 11, no. 10: 721. https://doi.org/10.3390/insects11100721
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.