
Multifunctionality of Jasmonic Acid Accumulation during Aphid Infestation in Altering the Plant Physiological Traits That Suppress the Plant Defenses in Wheat Cultivar XN979


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Insect Maintenance
2.2. Plant Growth Conditions
2.3. The Preference of S. miscanthi for Phytohormone-Treated Hosts
2.4. The Performance of S. miscanthi on Seedlings Treated with Phytohormones
2.5. Gene Expression Profiles in Wheat Plants after Phytohormone Treatments
2.6. Statistical Analyses
3. Results
3.1. Direct Foliage Treatment with Phytohormone Affected the Preference and Oviposition of S. miscanthi
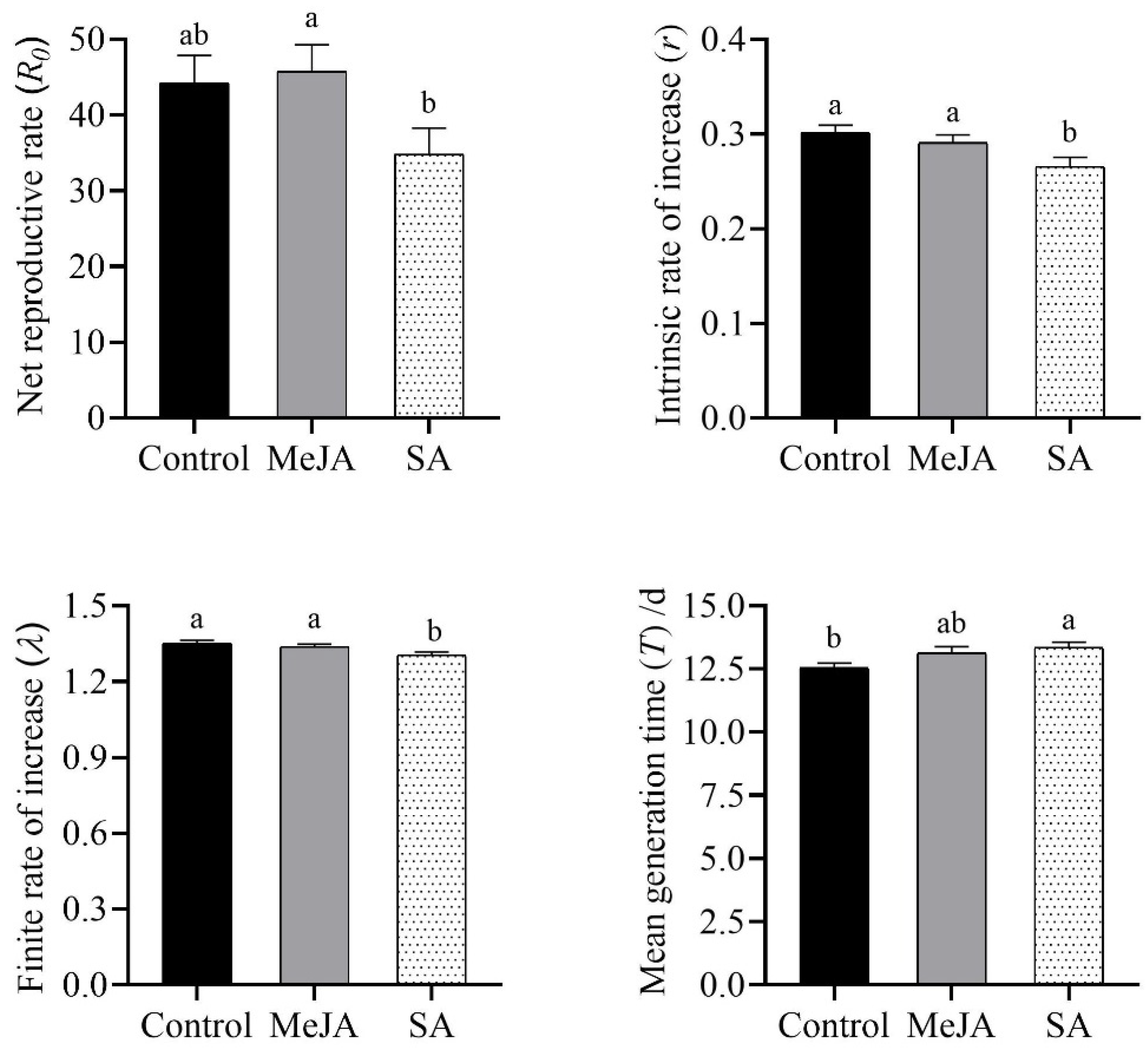
3.2. MeJA Application Weakly Exerted Positive Effects on the Performance of S. miscanthi
3.3. MeJA Treatments Significantly Elevated the Marker Genes Involved in JA Signaling in Plants
3.4. MeJA Treatments Significantly Suppressed the Marker Genes Involved in SA Signaling, While Elevating Tryptamine Biosynthesis Gene
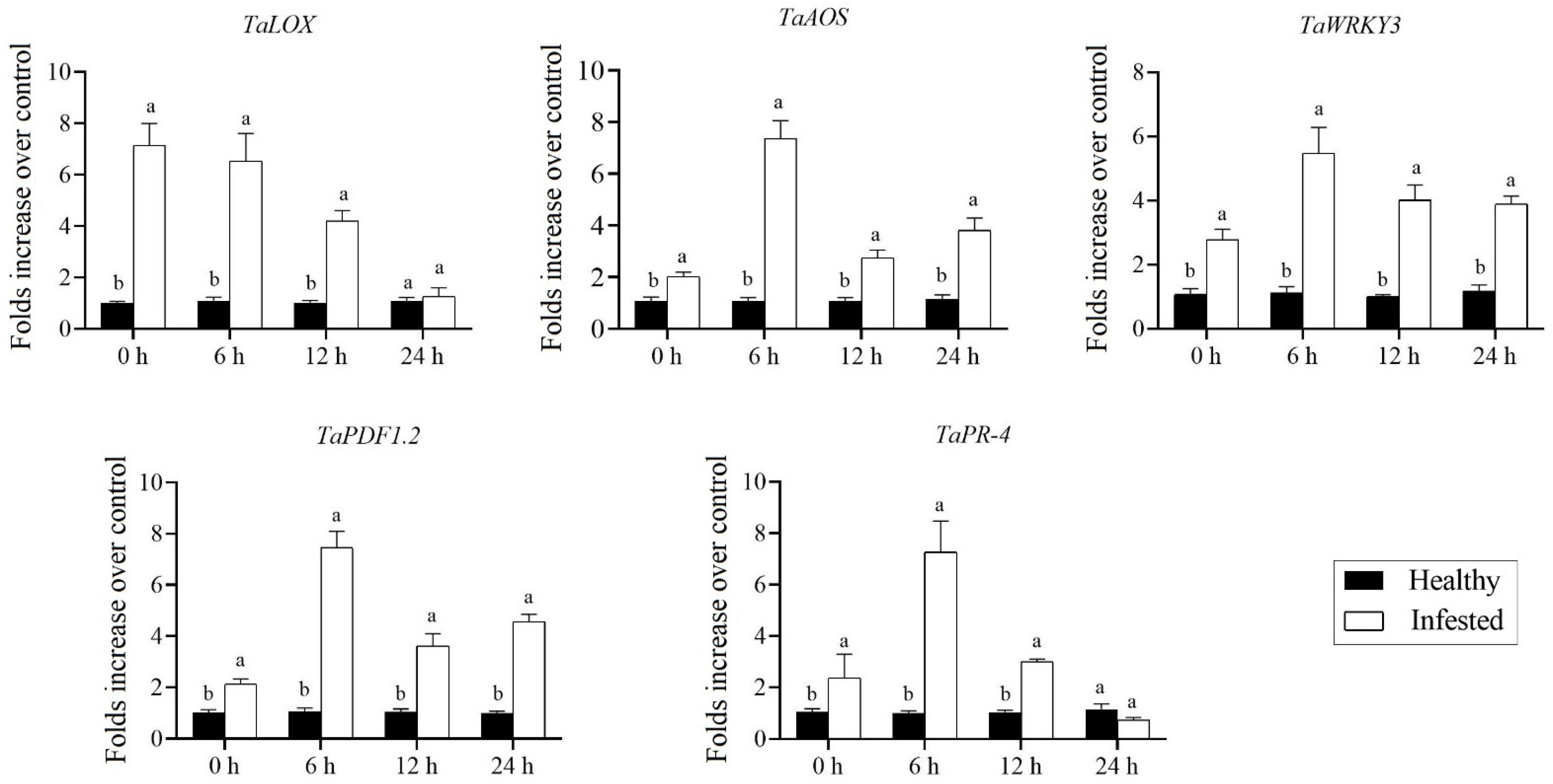
3.5. Aphid Infestation Significantly Triggered the Expression of SA- and JA-Dependent Response Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bouvet, L.; Holdgate, S.; James, L.; Thomas, J.; Mackay, I.J.; Cockram, J. The evolving battle between yellow rust and wheat: Implications for global food security. Theor. Appl. Genet. 2022, 135, 741–753. [Google Scholar] [CrossRef]
- Luo, K.; Zhao, H.; Wang, X.; Kang, Z. Prevalent pest management strategies for grain aphids: Opportunities and challenges. Front. Plant Sci. 2022, 12, 3252. [Google Scholar] [CrossRef]
- Sun, J.; Li, Q.; Tan, X.; Fan, J.; Zhang, Y.; Qin, Y.; Francis, F.; Chen, J. Population genetic structure of Sitobion miscanthi in China. J. Integr. Agric. 2022, 21, 178–187. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Sabri, A.; Vandermoten, S.; Leroy, P.D.; Haubruge, E.; Hance, T.; Thonart, P.; De Pauw, E.; Francis, F. Proteomic investigation of aphid honeydew reveals an unexpected diversity of proteins. PLoS ONE 2013, 8, e74656. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, R.; Atamian, H.S.; Shen, Z.; Briggs, S.P.; Kaloshian, I. GroEL from the endosymbiont Buchnera aphidicola betrays the aphid by triggering plant defense. Proc. Natl. Acad. Sci. USA 2014, 111, 8919–8924. [Google Scholar] [CrossRef]
- Jaouannet, M.L.; Rodriguez, P.A.; Thorpe, P.; Lenoir, C.J.G.; MacLeod, R.; Escudero-Martinez, C.; Bos, J.I.B. Plant immunity in plant-aphid interactions. Front. Plant Sci. 2014, 5, 663. [Google Scholar] [CrossRef] [Green Version]
- Rajendra, B.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Gill, R.S.; Gupta, A.K.; Taggar, G.K.; Taggar, M.S. Role of oxidative enzymes in plant defenses against insect herbivory. Acta Phytopathol. Entomol. Hung. 2010, 45, 277–290. [Google Scholar] [CrossRef]
- Verhage, A.; Van Wees, S.C.M.; Pieterse, C.M.J. Plant immunity: It’s the hormones talking, but what do they say? Plant Physiol. 2010, 154, 536–540. [Google Scholar] [CrossRef] [Green Version]
- Qi, P.; Balcerzak, M.; Rocheleau, H.; Leung, W.; Wei, Y.; Zheng, Y.; Ouellet, T. Jasmonic acid and abscisic acid play important roles in host-pathogen interaction between Fusarium graminearum and wheat during the early stages of Fusarium head blight. Physiol. Mol. Plant Pathol. 2016, 93, 39–48. [Google Scholar] [CrossRef]
- Walling, L.L. The Myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef]
- Moran, P.J.; Thompson, G.A. Molecular responses to aphid feeding in Arabidopsis in relation to plant defense pathways. Plant Physiol. 2001, 125, 1074–1085. [Google Scholar] [CrossRef] [Green Version]
- Rayapuram, C.; Baldwin, I. Increased SA in NPR1-silenced plants antagonizes JA and JA-dependent direct and indirect defenses in herbivore-attacked Nicotiana attenuata in nature. Plant J. 2007, 52, 700–715. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Chen, J.L.; Cheng, D.F.; Sun, J.R.; Liu, Y.; Tian, Z. Biochemical and molecular characterizations of Sitobion avenae-induced wheat defense responses. Crop. Prot. 2009, 28, 435–442. [Google Scholar] [CrossRef]
- Shivaji, R.; Camas, A.; Ankala, A.; Engelberth, J.; Tumlinson, J.H.; Williams, W.P.; Wilkinson, J.R.; Luthe, D.S. Plants on constant alert: Elevated levels of jasmonic acid and jasmonate-induced transcripts in caterpillar-resistant maize. J. Chem. Ecol. 2010, 36, 179–191. [Google Scholar] [CrossRef]
- Cao, H.; Wang, S.; Liu, T. Jasmonate- and salicylate-induced defenses in wheat affect host preference and probing behavior but not performance of the grain aphid, Sitobion avenae. Insect Sci. 2014, 21, 47–55. [Google Scholar] [CrossRef]
- Luo, K.; Yao, X.; Luo, C.; Hu, X.; Hu, Z.; Zhang, G.; Zhao, H. Previous aphid infestation induces different expression profiles of genes associated with hormone-dependent responses in near-isogenic winter wheat lines. J. Econ. Entomol. 2020, 113, 461–470. [Google Scholar] [CrossRef]
- Pandey, D.; Rajendran, S.R.C.K.; Gaur, M.; Sajeesh, P.K.; Kumar, A. Plant defense signaling and responses against necrotrophic fungal pathogens. J. Plant Growth Regul. 2016, 35, 1159–1174. [Google Scholar] [CrossRef]
- Shigenaga, A.M.; Argueso, C.T. No hormone to rule them all: Interactions of plant hormones during the responses of plants to pathogens. Semin. Cell Dev. Biol. 2016, 56, 174–189. [Google Scholar] [CrossRef]
- Xu, H.; Qian, L.; Wang, X.; Shao, R.; Hong, Y.; Liu, S.; Wang, X. A salivary effector enables whitefly to feed on host plants by eliciting salicylic acid-signaling pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Poelman, E.H.; Broekgaarden, C.; Van Loon, J.J.; Dicke, M. Early season herbivore differentially affects plant defence responses to subsequently colonizing herbivores and their abundance in the field. Mol. Ecol. 2008, 17, 3352–3365. [Google Scholar] [CrossRef]
- Snoeren, T.A.L.; Broekgaarden, C.; Dicke, M. Jasmonates differentially affect interconnected signal-transduction pathways of Pieris rapae-induced defenses in Arabidopsis thaliana. Insect Sci. 2011, 18, 249–258. [Google Scholar] [CrossRef]
- Luo, K.; Ouellet, T.; Zhao, H.; Wang, X.; Kang, Z. Wheat—Fusarium graminearum interactions under Sitobion avenae influence: From nutrients and hormone signals. Front. Nutr. 2021, 8, 703293. [Google Scholar] [CrossRef]
- Lu, H.; Luo, T.; Fu, H.; Wang, L.; Tan, Y.; Huang, J.; Wang, Q.; Ye, G.; Gatehouse, A.M.R.; Lou, Y.; et al. Resistance of rice to insect pests mediated by suppression of serotonin biosynthesis. Nat. Plants 2018, 4, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Brauer, E.K.; Rocheleau, H.; Balcerzak, M.; Pan, Y.; Fauteux, F.; Liu, Z.; Wang, L.; Zheng, W.; Ouellet, T. Transcriptional and hormonal profiling of Fusarium graminearum-infected wheat reveals an association between auxin and susceptibility. Physiol. Mol. Plant Pathol. 2019, 107, 33–39. [Google Scholar] [CrossRef]
- Su, P.; Zhao, L.; Li, W.; Zhao, J.; Yan, J.; Ma, X.; Li, A.; Wang, H.; Kong, L. Integrated metabolo-transcriptomics and functional characterization reveals that the wheat auxin receptor TIR1 negatively regulates defense against Fusarium graminearum. J. Integr. Plant Biol. 2021, 63, 340–352. [Google Scholar] [CrossRef]
- Tsavkelova, E.; Oeser, B.; Oren-Young, L.; Israeli, M.; Sasson, Y.; Tudzynski, B.; Sharon, A. Identification and functional characterization of indole-3-acetamide-mediated IAA biosynthesis in plant-associated Fusarium species. Fungal Genet. Biol. 2012, 49, 48–57. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Plants-Entwicklungsstadien von Pflanzen-Estadios de las Plantas-Développement des Plantes. In BBCH-Monograph; Blackwell Wissenschafsverlag: Berlin, Germany, 1997; p. 622. [Google Scholar]
- Latini, A.; Fiorani, F.; Galeffi, P.; Cantale, C.; Bevivino, A.; Jablonowski, N.D. Phenotyping of different Italian durum wheat varieties in early growth stage with the addition of pure or digestate-activated biochars. Front. Plant Sci. 2021, 12, 782072. [Google Scholar] [CrossRef]
- Wei, M.; Chi, H.; Guo, Y.; Li, X.; Zhao, L.; Ma, R. Demography of Cacopsylla chinensis (Hemiptera: Psyllidae) reared on four cultivars of Pyrus bretschneideri (Rosales: Rosaceae) and P. communis pears with estimations of confidence intervals of specific life table statistics. J. Econ. Entomol. 2020, 113, 2343–2353. [Google Scholar] [CrossRef]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Worrall, D.; Holroyd, G.H.; Moore, J.P.; Glowacz, M.; Croft, P.; Taylor, J.E.; Paul, N.D.; Roberts, M.R. Treating seeds with activators of plant defence generates long-lasting priming of resistance to pests and pathogens. New Phytol. 2012, 193, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Zhang, M.; Yang, K.; Zheng, C. Salicylic acid-primed defence response in octoploid strawberry ‘Benihoppe’ leaves induces resistance against Podosphaera aphanis through enhanced accumulation of proanthocyanidins and upregulation of pathogenesis-related genes. BMC Plant Biol. 2020, 20, 149. [Google Scholar] [CrossRef] [Green Version]
- Paranidharan, V.; Abu-Nada, Y.; Hamzehzarghani, H.; Kushalappa, A.C.; Mamer, O.; Dion, Y.; Rioux, S.; Comeau, A.; Choiniere, L. Resistance-related metabolites in wheat against Fusarium graminearum and the virulence factor deoxynivalenol (DON). Botany 2008, 86, 1168–1179. [Google Scholar] [CrossRef]
- Kumaraswamy, G.K.; Bollina, V.; Kushalappa, A.C.; Choo, T.M.; Dion, Y.; Rioux, S.; Mamer, O.; Faubert, D. Metabolomics technology to phenotype resistance in barley against Gibberella zeae. Eur. J. Plant Pathol. 2011, 130, 29–43. [Google Scholar] [CrossRef]
- Pasquet, J.; Chaouch, S.; Macadré, C.; Balzergue, S.; Huguet, S.; Martin-Magniette, M.; Bellvert, F.; Deguercy, X.; Thareau, V.; Heintz, D.; et al. Differential gene expression and metabolomic analyses of Brachypodium distachyon infected by deoxynivalenol producing and non-producing strains of Fusarium graminearum. BMC Genom. 2014, 15, 629. [Google Scholar] [CrossRef] [Green Version]
- Luo, K.; DesRoches, C.; Johnston, A.; Harris, L.; Zhao, H.; Ouellet, T. Multiple metabolic pathways for metabolism of L-tryptophan in Fusarium graminearum. Can. J. Microbiol. 2017, 63, 921–927. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Zhao, D.; Zhang, C.; Wu, H.; Li, Q.; Gu, M.; Sun, S.S.; Liu, Q. A connection between lysine and serotonin metabolism in rice endosperm. Plant Physiol. 2018, 176, 1965–1980. [Google Scholar] [CrossRef] [Green Version]
- Luo, K.; Rocheleau, H.; Qi, P.; Zheng, Y.; Zhao, H.; Ouellet, T. Indole-3-acetic acid in Fusarium graminearum: Identification of biosynthetic pathways and characterization of physiological effects. Fungal Biol. 2016, 120, 1135–1145. [Google Scholar] [CrossRef]
- Fu, J.; Wang, S. Insights into Auxin signaling in plant-pathogen interactions. Front. Plant Sci. 2011, 2, 74. [Google Scholar] [CrossRef] [Green Version]
- Weeraddana, C.D.S.; Manolii, V.P.; Strelkov, S.E.; de la Mata, A.P.; Harynuk, J.J.; Evenden, M.L. Infection of canola by the root pathogen Plasmodiophora brassicae increases resistance to aboveground herbivory by bertha armyworm, Mamestra configurata Walker (Lepidoptera: Noctuidae). Plant Sci. 2020, 300, 110625. [Google Scholar] [CrossRef]
- Koch, A.; Kumar, N.; Weber, L.; Keller, H.; Imani, J.; Kogel, K. Host-induced gene silencing of cytochrome P450 lanosterol C14α-demethylase-encoding genes confers strong resistance to Fusarium species. Proc. Natl. Acad. Sci. USA 2013, 110, 19324–19329. [Google Scholar] [CrossRef]
- Liu, S.; Geng, S.; Li, A.; Mao, Y.; Mao, L. RNAi technology for plant protection and its application in wheat. aBiotech 2021, 2, 365–374. [Google Scholar] [CrossRef]
- Cao, H.H.; Pan, M.Z.; Liu, H.R.; Wang, S.H.; Liu, T.X. Antibiosis and tolerance but not antixenosis to the grain aphid, Sitobion avenae (Hemiptera: Aphididae), are essential mechanisms of resistance in a wheat cultivar. Bull. Entomol. Res. 2015, 105, 448–455. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Treatments | ||
|---|---|---|---|
| Control | MeJA | SA | |
| Adult preoviposition period, (d) | 7.52 ± 0.11 c | 7.96 ± 0.14 b | 8.57 ± 0.24 a |
| Total preoviposition period, (d) | 0.60 ± 0.10 b | 0.89 ± 0.08 a | 1.25 ± 0.25 a |
| Oviposition days, (d) | 13.64 ± 0.90 a | 14.57 ± 0.63 a | 12.46 ± 0.77 a |
| Oviposition period, (d) | 14.28 ± 0.99 a | 15.57 ± 0.62 a | 13.18 ± 0.73 a |
| Daily larvae production during oviposition period | 3.30 ± 0.19 a | 3.13 ± 0.17 a | 2.75 ± 0.18 a |
| Fecundity (F) | 46.01 ± 3.32 ab | 49.03 ± 2.90 a | 37.30 ± 3.23 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, X.; Xu, S.; Guo, J.; Hu, J.; He, D.; Jia, L.; Shang, H.; Li, G.; Luo, K. Multifunctionality of Jasmonic Acid Accumulation during Aphid Infestation in Altering the Plant Physiological Traits That Suppress the Plant Defenses in Wheat Cultivar XN979. Insects 2023, 14, 622. https://doi.org/10.3390/insects14070622
Yan X, Xu S, Guo J, Hu J, He D, Jia L, Shang H, Li G, Luo K. Multifunctionality of Jasmonic Acid Accumulation during Aphid Infestation in Altering the Plant Physiological Traits That Suppress the Plant Defenses in Wheat Cultivar XN979. Insects. 2023; 14(7):622. https://doi.org/10.3390/insects14070622
Chicago/Turabian StyleYan, Xia, Shicai Xu, Jiao Guo, Jiazhen Hu, Dejia He, Li Jia, Huanzhang Shang, Guangwei Li, and Kun Luo. 2023. "Multifunctionality of Jasmonic Acid Accumulation during Aphid Infestation in Altering the Plant Physiological Traits That Suppress the Plant Defenses in Wheat Cultivar XN979" Insects 14, no. 7: 622. https://doi.org/10.3390/insects14070622