
Analysis of the Propionate Metabolism in Bacillus subtilis during 3-Indolacetic Production
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Media Cultivation
2.2. Bioreactor Cultivation
2.3. Biomass and Metabolite Quantification
2.4. Genome-Scale Metabolic Model and Flux Calculations
2.5. Constrain-Based Analyses: Robustness, Gene Deletion, Phenotypic Phase Plane and OptKnock
2.6. Statistical Analysis and Data Chart
3. Results
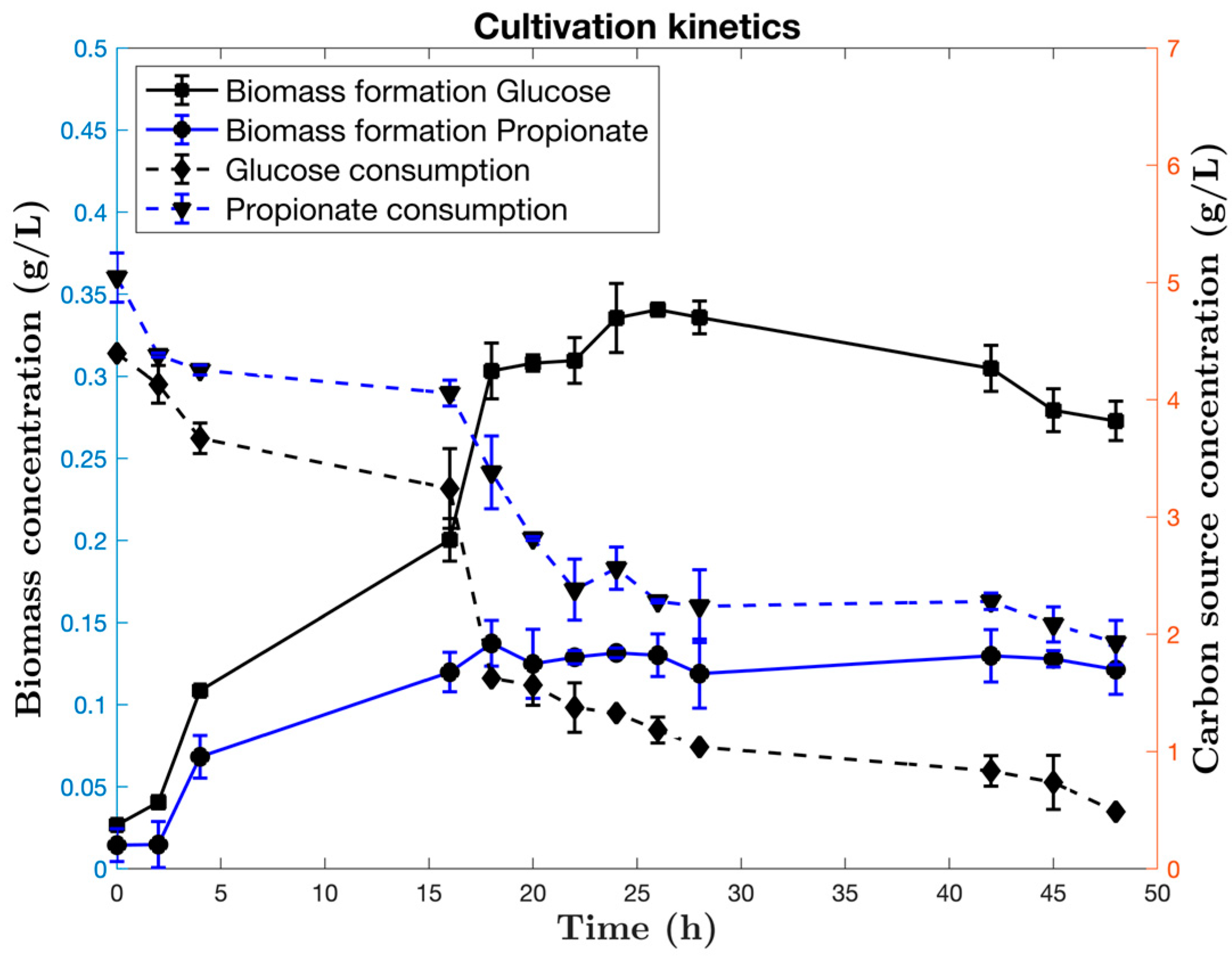
3.1. Bioreactor Cultivations
3.1.1. Cultivation Kinetics
3.1.2. IAA Kinetics
3.1.3. Kinetic Parameters
3.2. Flux Distributions
3.3. Propionate Uptake Pathway
3.4. Analysis of Metabolic Capabilities
3.4.1. Simple Robustness Analysis
3.4.2. Double Robustness Analysis
3.4.3. Single Gene Deletion
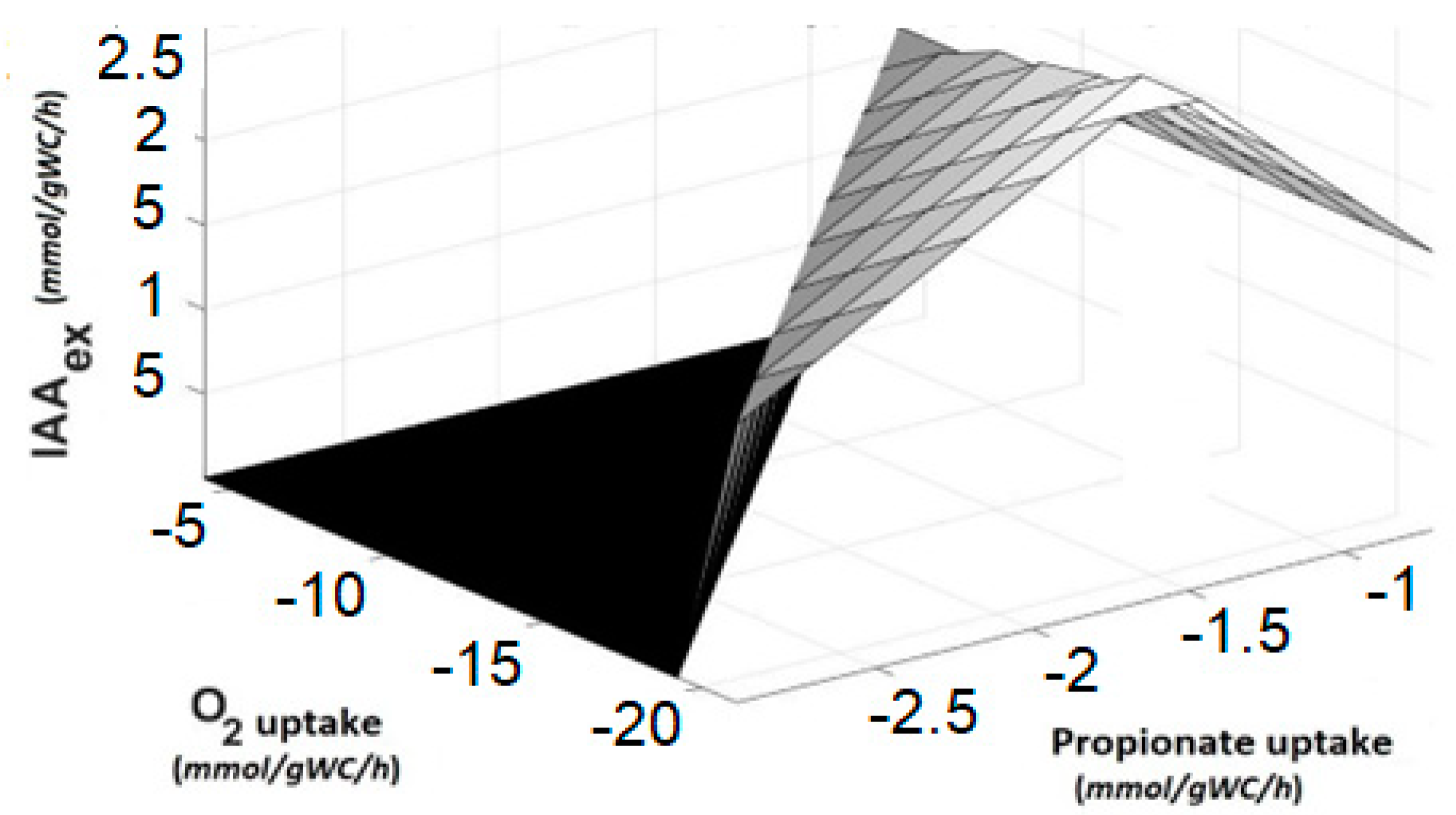
3.4.4. Phenotype Phase Plane Analysis
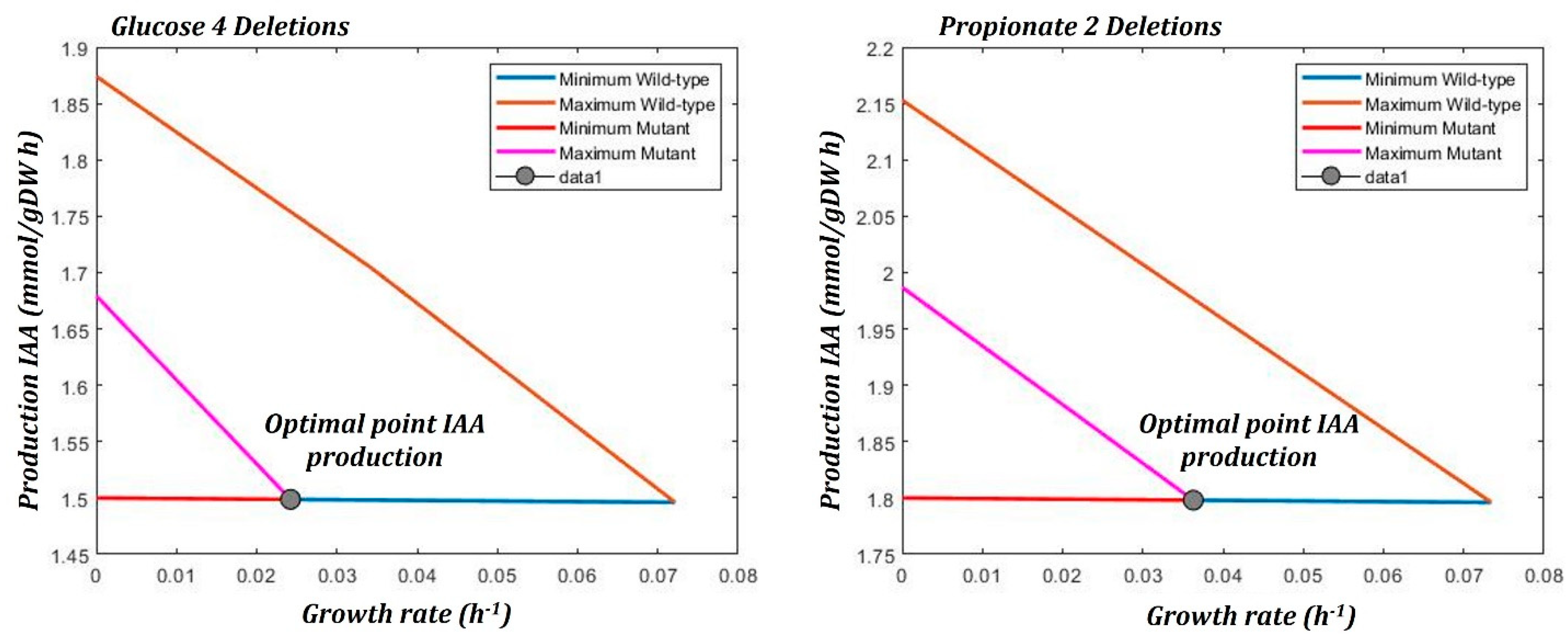
3.4.5. Bilevel Optimization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| glucose –> or propionate –> |
| g6p <=> f6p |
| atp + f6p <=> adp + f1,6bf |
| f1,6bf <=> g3p + dhp |
| g3p + nad + pi <=> 1,3dpg + h + nadh |
| atp + h + 3pg <=> adp + 1,3dpg |
| 2pg <=> 3pg |
| 3pg <=> h2o + pep |
| adp + pep <=> atp + pyr |
| pyr + Coa + nad <=> acoa + nadh + co_2 |
| ACoa + h_2o + oaa <=> h + coa + cit |
| cit <=> acon-C + h_2o |
| acon-C + h_2o <=> icit |
| icit + nadp <=> akg + co_2 + nadph |
| akg + Coa + nadh –> co_2 + nadh + succoa |
| atp + Coa + succ <=> adp + pi + succoa |
| q8 + succ –> fum + q8h2 |
| fum + h_2o <=> mal |
| mal + nad => h + nadh + oaa |
| atp + oaa –> adp + co_2 + pep |
| atp +pyr <=> adp + p_i + h + oaa |
| mal-L + nad –> co_2 + nadh + pyr |
| mal-L + nadp –> co_2 + nadph + pyr |
| g6p + nadp <=> 6pgl + h + nadph |
| 6pgc + nadp –> co_2 + nadph + ru5p |
| ru5p <==> xu5p |
| r5p <=> ru5p |
| e4p + xu5p <=> f6p + g3p |
| g3p + sh7p <=> f6p + e4p |
| ser + igp <=> g3p + h_2o + trp |
| tryptophan –> |
| trp <=> iam |
| h_2o + iam <=> nh_4 + iaa |
Appendix B
| Locus | Flux | Protein |
|---|---|---|
| BSU13350 | 2.18 | tryptophan synthase alpha chain |
| BSU13930 | 2.18 | undespore photoproduct lyase |
| BSU21920 | 2.15 | processive diacylglycerol glucosyltransferase |
| BSU35760 | 1.77 | glycerol phosphate glycerophosphotransferase |
| BSU35660 | 1.77 | UDP-N-acetylglucosamine 2-epimerase |
| BSU35700 | 1.77 | ABC transporter |
| BSU35710 | 1.77 | ABC transporter |
| BSU3572 | 1.75 | glycerol-phosphotransferase |
| BSU35530 | 1.75 | GlcNAc-1-phosphate transferase |
| BSU35750 | 1.74 | UDP-N-acetyl-D-mannosamine transferase |
| BSU35730 | 1.72 | polyglycerol phosphate GlcNAc-1-phosphate transferase |
| BSU35750 | 1.74 | UDP-N-acetyl-D-mannosamine transferase |
| BSU35730 | 1.72 | polyglycerol phosphate glucosyltransferase |
References
- Brumos, J.; Robles, L.M.; Yun, J.; Vu, T.C.; Jackson, S.; Alonso, J.M.; Stepanova, A.N. Local Auxin Biosynthesis Is a Key Regulator of Plant Development. Dev. Cell 2018, 47, 306–318.e5. [Google Scholar] [CrossRef] [Green Version]
- Grossmann, K. Auxin Herbicides: Current Status of Mechanism and Mode of Action. Pest. Manag. Sci. 2010, 66, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Dobbelaere, S.; Vanderleyden, J.; Okon, Y. Plant Growth-Promoting Effects of Diazotrophs in the Rhizosphere. Crit. Rev. Plant Sci. 2003, 22, 107–149. [Google Scholar] [CrossRef]
- Fischer, C.R.; Klein-Marcuschamer, D.; Stephanopoulos, G. Selection and Optimization of Microbial Hosts for Biofuels Production. Metab. Eng. 2008, 10, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Buffing, M.F.; Link, H.; Christodoulou, D.; Sauer, U. Capacity for Instantaneous Catabolism of Preferred and Non-Preferred Carbon Sources in Escherichia coli and Bacillus subtilis. Sci. Rep. 2018, 8, 11760. [Google Scholar] [CrossRef] [Green Version]
- Sridevi, M.; Yadav, N.C.S.; Mallaiah, K.V. Production of Indole-Acetic-Acid by Rhizobium Isolates from Crotalaria Species. Res. J. Microbiol. 2008, 3, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Halmschlag, B.; Putri, S.P.; Fukusaki, E.; Blank, L.M. Poly-γ-Glutamic Acid Production by Bacillus subtilis 168 Using Glucose as the Sole Carbon Source: A Metabolomic Analysis. J. Biosci. Bioeng. 2020, 130, 272–282. [Google Scholar] [CrossRef]
- Rainey, F.; Kämpfer, P.; Trujillo, M.; Chun, J.; DeVos, P. Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; Wiley: Hoboken, NJ, USA, 2015; Volume 410. [Google Scholar]
- Zhao, J.; Shimizu, K. Metabolic Flux Analysis of Escherichia coli K12 Grown on 13C-Labeled Acetate and Glucose Using GC-MS and Powerful Flux Calculation Method. J. Biotechnol. 2003, 101, 101–117. [Google Scholar] [CrossRef]
- Araújo, F.F.; Henning, A.A.; Hungria, M. Phytohormones and Antibiotics Produced by Bacillus subtilis and Their Effects on Seed Pathogenic Fungi and on Soybean Root Development. World J. Microbiol. Biotechnol. 2005, 21, 1639–1645. [Google Scholar] [CrossRef]
- Zaidi, S.; Usmani, S.; Singh, B.R.; Musarrat, J. Significance of Bacillus subtilis Strain SJ-101 as a Bioinoculant for Concurrent Plant Growth Promotion and Nickel Accumulation in Brassica Juncea. Chemosphere 2006, 64, 991–997. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, D.; Qi, G.; Mao, Z.; Hu, X.; Du, B.; Liu, K.; Ding, Y. Effects of Bacillus velezensis FKM10 for Promoting the Growth of Malus hupehensis Rehd. and Inhibiting Fusarium verticillioides. Front. Microbiol. 2020, 10, 2889. [Google Scholar] [CrossRef]
- Escobar Diaz, P.A.; Gil, O.J.A.; Barbosa, C.H.; Desoignies, N.; Rigobelo, E.C. Aspergillus spp. and Bacillus spp. as Growth Promoters in Cotton Plants Under Greenhouse Conditions. Front. Sustain. Food Syst. 2021, 5, 709267. [Google Scholar] [CrossRef]
- Myo, E.M.; Ge, B.; Ma, J.; Cui, H.; Liu, B.; Shi, L.; Jiang, M.; Zhang, K. Indole-3-Acetic Acid Production by Streptomyces fradiae NKZ-259 and Its Formulation to Enhance Plant Growth. BMC Microbiol. 2019, 19, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Yang, J.; Shen, P.; Li, Q.; Wu, W.; Jiang, X.; Qin, L.; Huang, J.; Cao, X.; Qi, F. High-Level Production of Indole-3-Acetic Acid in the Metabolically Engineered Escherichia coli. J. Agric. Food Chem. 2021, 69, 1916–1924. [Google Scholar] [CrossRef] [PubMed]
- Bunsangiam, S.; Thongpae, N.; Limtong, S.; Srisuk, N. Large Scale Production of Indole-3-Acetic Acid and Evaluation of the Inhibitory Effect of Indole-3-Acetic Acid on Weed Growth. Sci. Rep. 2021, 11, 13094. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Kim, G.B.; Kim, W.J.; Kim, H.U.; Lee, S.Y. Current Status and Applications of Genome-Scale Metabolic Models. Genome Biol. 2019, 20, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orth, J.D.; Thiele, I.; Palsson, B.O. What Is Flux Balance Analysis? Nat. Biotechnol. 2010, 28, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, S.N.; Olivier, B.G.; Molenaar, D.; Teusink, B. A Systematic Assessment of Current Genome-Scale Metabolic Reconstruction Tools. Genome Biol. 2019, 20, 158. [Google Scholar] [CrossRef] [Green Version]
- Lularevic, M.; Racher, A.J.; Jaques, C.; Kiparissides, A. Improving the Accuracy of Flux Balance Analysis through the Implementation of Carbon Availability Constraints for Intracellular Reactions. Biotechnol. Bioeng. 2019, 116, 2339–2352. [Google Scholar] [CrossRef]
- Burgard, A.P.; Pharkya, P.; Maranas, C.D. Optknock: A Bilevel Programming Framework for Identifying Gene Knockout Strategies for Microbial Strain Optimization. Biotechnol. Bioeng. 2003, 84, 647–657. [Google Scholar] [CrossRef]
- He, M.; Wen, J.; Yin, Y.; Wang, P. Metabolic Engineering of Bacillus subtilis Based on Genome-Scale Metabolic Model to Promote Fengycin Production. 3 Biotech 2021, 11, 448. [Google Scholar] [CrossRef] [PubMed]
- Costanza, J.; Carapezza, G.; Angione, C.; Liò, P.; Nicosia, G. Multi-Objective Optimisation, Sensitivity and Robustness Analysis in FBA Modelling. In Lecture Notes in Computer Science (including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics); Springer: Heidelberg, Germany, 2012; Volume 7605. [Google Scholar]
- Karimi, E.; Geslain, E.; Belcour, A.; Frioux, C.; Aïte, M.; Siegel, A.; Corre, E.; Dittami, S.M. Robustness Analysis of Metabolic Predictions in Algal Microbial Communities Based on Different Annotation Pipelines. PeerJ 2021, 9, e11344. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.S.; Palsson, B.O. Robustness Analysis of the Escherichia coli Metabolic Network. Biotechnol. Prog. 2000, 16, 927–939. [Google Scholar] [CrossRef]
- Edwards, J.S.; Ramakrishna, R.; Palsson, B.O. Characterizing the Metabolic Phenotype: A Phenotype Phase Plane Analysis. Biotechnol. Bioeng. 2002, 77, 27–36. [Google Scholar] [CrossRef]
- Iman, M.; Sobati, T.; Panahi, Y.; Mobasheri, M. Systems Biology Approach to Bioremediation of Nitroaromatics: Constraint-Based Analysis of 2,4,6-Trinitrotoluene Biotransformation by Escherichia coli. Molecules 2017, 22, 1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salleh, A.H.M.; Mohamad, M.S.; Deris, S.; Omatu, S.; Fdez-Riverola, F.; Corchado, J.M. Gene Knockout Identification for Metabolite Production Improvement Using a Hybrid of Genetic Ant Colony Optimization and Flux Balance Analysis. Biotechnol. Bioprocess Eng. 2015, 20, 685–693. [Google Scholar] [CrossRef]
- Heirendt, L.; Arreckx, S.; Pfau, T.; Mendoza, S.N.; Richelle, A.; Heinken, A.; Haraldsdóttir, H.S.; Wachowiak, J.; Keating, S.M.; Vlasov, V.; et al. Creation and Analysis of Biochemical Constraint-Based Models Using the COBRA Toolbox v.3.0. Nat. Protoc. 2019, 14, 639–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo Alfonso, F.; Vigueras-Ramírez, G.; Rosales-Colunga, L.M.; del Monte-Martínez, A.; Olivares Hernández, R. Propionate as the Preferred Carbon Source to Produce 3-Indoleacetic Acid In B. subtilis: Comparative Flux Analysis Using Five Carbon Sources. Mol. Omics 2021, 17, 554–564. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A Critical Examination of the Specificity of the Salkowski Reagent for Indolic Compounds Produced by Phytopathogenic Bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [Green Version]
- Hao, T.; Han, B.; Ma, H.; Fu, J.; Wang, H.; Wang, Z.; Tang, B.; Chen, T.; Zhao, X. In Silico Metabolic Engineering of Bacillus Subtilis for Improved Production of Riboflavin, Egl-237, (R,R)-2,3-Butanediol and Isobutanol. Mol. Biosyst. 2013, 9, 2034–2044. [Google Scholar] [CrossRef]
- Streif, S.; Kim, K.K.K.; Rumschinski, P.; Kishida, M.; Shen, D.E.; Findeisen, R.; Braatz, R.D. Robustness Analysis, Prediction, and Estimation for Uncertain Biochemical Networks: An Overview. J. Process Control 2016, 42, 14–34. [Google Scholar] [CrossRef]
- Anand, S.; Mukherjee, K.; Padmanabhan, P. An Insight to Flux-Balance Analysis for Biochemical Networks. Biotechnol. Genet. Eng. Rev. 2020, 36, 32–55. [Google Scholar] [CrossRef]
- Price, N.D.; Reed, J.L.; Palsson, B.Ø. Genome-Scale Models of Microbial Cells: Evaluating the Consequences of Constraints. Nat. Rev. Microbiol. 2004, 2, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Pharkya, P.; Burgard, A.P.; Maranas, C.D. Exploring the overproduction of amino acids using the bilevel optimization framework OptKnock. Biotechnol. Bioeng. 2003, 84, 887–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKight, P.E.; Najab, J. Kruskal-Wallis Test. In The Corsini Encyclopedia of Psychology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010. [Google Scholar]
- Anderson, D.R.; Sweeney, D.J.; Williams, T.A.; Camm, J.D.; Cochran, J.J. Modern Business Statistics with Microsoft Excel; Cengage Learning: Boston, MA, USA, 2020; ISBN 0357131525. [Google Scholar]
- Doyle, J.S.H. Towards the Stereoselective Syntheses of 2-Methylcitrate and 2-Methylisocitrate and Their Use in Studying the Methylcitric Acid Cycle in Bacillus subtilis; The University of North Carolina at Greensboro: Greensboro, NC, USA, 2018; ISBN 0438087801. [Google Scholar]
- Sander, T.; Farke, N.; Diehl, C.; Kuntz, M.; Glatter, T.; Link, H. Allosteric Feedback Inhibition Enables Robust Amino Acid Biosynthesis in E. Coli by Enforcing Enzyme Overabundance. Cell Syst. 2019, 8, 66–75.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Ehrlich, S.D.; Albertini, A.; Amati, G.; Andersen, K.K.; Arnaud, M.; Asai, K.; Ashikaga, S.; Aymerich, S.; Bessieres, P.; et al. Essential Bacillus subtilis Genes. Proc. Natl. Acad. Sci. USA 2003, 100, 4678–4683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, B.-M.; Kritikos, G.; Farelli, J.D.; Todor, H.; Tong, K.; Kimsey, H.; Wapinski, I.; Galardini, M.; Cabal, A.; Peters, J.M.; et al. Construction and Analysis of Two Genome-Scale Deletion Libraries for Bacillus subtilis. Cell Syst. 2017, 4, 291–305.e7. [Google Scholar] [CrossRef]
- Michna, R.H.; Zhu, B.; Mäder, U.; Stülke, J. SubtiWiki 2.0—An Integrated Database for the Model Organism Bacillus subtilis. Nucleic Acids Res. 2016, 44, D654–D662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boele, J.; Olivier, B.G.; Teusink, B. FAME, the Flux Analysis and Modeling Environment. BMC Syst. Biol. 2012, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG Mapping Tools for Uncovering Hidden Features in Biological Data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A. UniProt: A Worldwide Hub of Protein Knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- Ehrenshaft, M.; Deterding, L.J.; Mason, R.P. Tripping up Trp: Modification of Protein Tryptophan Residues by Reactive Oxygen Species, Modes of Detection, and Biological Consequences. Free Radic. Biol. Med. 2015, 89, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Lei, P.; Mohsin, A.; Liu, X.; Huang, M.; Li, L.; Hu, J.; Hang, H.; Zhuang, Y.; Guo, M. Mixomics Analysis of Bacillus Subtilis: Effect of Oxygen Availability on Riboflavin Production. Microb. Cell Fact. 2017, 16, 150. [Google Scholar] [CrossRef] [PubMed]
- Junne, S.; Klingner, A.; Kabisch, J.; Schweder, T.; Neubauer, P. A Two-Compartment Bioreactor System Made of Commercial Parts for Bioprocess Scale-down Studies: Impact of Oscillations on Bacillus subtilis Fed-Batch Cultivations. Biotechnol. J. 2011, 6, 1009–1017. [Google Scholar] [CrossRef]
- Sivasakthi, S.; Kanchana, D.; Usharani, G.; Saranraj, P. Production of plant growth promoting substance by Pseudomonas fluorescens and Bacillus subtilis isolates from paddy rhizosphere soil of Cuddalore District, Tamil Nadu, India. Int. J. Microbiol. Res. 2013, 4, 227–233. [Google Scholar]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-Acetic Acid in Plant–Microbe Interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Defez, R.; Andreozzi, A.; Bianco, C. The Overproduction of Indole-3-Acetic Acid (IAA) in Endophytes Upregulates Nitrogen Fixation in Both Bacterial Cultures and Inoculated Rice Plants. Microb. Ecol. 2017, 74, 441–452. [Google Scholar] [CrossRef]
- Niu, H.; Li, R.; Liang, Q.; Qi, Q.; Li, Q.; Gu, P. Metabolic Engineering for Improving L-tryptophan Production in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2019, 46, 55–65. [Google Scholar] [CrossRef]
- Zuo, S.; Xiao, J.; Zhang, Y.; Zhang, X.; Nomura, C.T.; Chen, S.; Wang, Q. Rational Design and Medium Optimization for Shikimate Production in Recombinant Bacillus licheniformis Strains. Process Biochem. 2018, 66, 19–27. [Google Scholar] [CrossRef]









| Parameter | Glucose | Propionate |
|---|---|---|
| μexp | 0.15 ± 0.06 | 0.14 ± 0.02 |
| μstat | 0.07 ± 0.12 | 0.06 ± 0.04 |
| qs,exp | 2.15 ± 0.08 | 4.18 ± 0.04 |
| qs,stat | 1.2 ± 0.03 | 2.85 ± 0.01 |
| qs,Trp | 1.5 ± 0.05 | 1.8 ± 0.02 |
| qp,IAA | 1.33 ± 0.02 | 1.92 ± 0.07 |
| Yx/s | 0.324 ± 0.05 | 0.219 ± 0.08 |
| Yx/(s+Trp) | 0.134 ± 0.11 | 0.094 ± 0.06 |
| Yp/(s+Trp) | 0.446 ± 0.03 | 0.524 ± 0.04 |
| Yp/Trp | 0.761 ± 0.04 | 0.915 ± 0.03 |
| GLUCOSE | Max ⱱbio | |||
|---|---|---|---|---|
| No. Del | Knockouts | Enzyme | Biomass (h−1) | IAA (mmol/gDW h) |
| 0 | “Wild type” | ------- | 0.1 | 1.496 |
| 2 | 1. FDP <=> DHAP + G3P 2. MAL <=> FUM + H2O | Fructose bisphosphate aldolase (fbaA) Fumarase (citG) | 0.07 | 1.4992 |
| 3 | 1. 2PG <=> H2O + PEP 2. MAL <=> FUM + H2O 3. G6P <=> F6P | Enolase (eno) Fumarase (citG) Glucose-6-phosphate isomerase (pgi) | 0.04 | 1.51 |
| 4 | 1. MAL + NAD <=> OAA + H + NADH 2. RU5P <=> XU5P 3. AKG + NAD + CoA<=> SUC-CoA + CO2 + NADH 4. F6P + ATP -> FDP + ADP | Malate dehydrogenase (mdh) Ribulose-5-phosphate epimerase (rpe) α-Ketoglutarate dehydrogenase (odhB) Phosphofructokinase (pfkA) | 0.04 | 1.61 |
| PROPIONATE | Max ⱱbio | |||
| No. Del | Knockouts | Enzyme | Biomass (h−1) | IAA (mmol/gDW h) |
| 0 | “Wild type” | ------- | 0.1 | 1.796 |
| 2 | 1. MAL + NAD <=> OAA + H + NADH 2. SUC-CoA + GDP + HPO4 <=> SUC + GTP + CoA | Malate dehydrogenase (mdh) Succinyl-CoA synthetase (sucCD) | 0.06 | 1.798 |
| 3 | 1. MAL + NAD <=> OAA + H + NADH 2. 2PG <=> H2O + PEP 3. AKG + NAD + CoA<=> SUC-CoA + CO2 + NADH | Malate dehydrogenase (mdh) Enolase (eno) α-Ketoglutarate dehydrogenase (odhB) | 0.03 | 1.798 |
| 5 | 1. AKG + ALA <=> GLU + PYR 2. MAL + NAD <=> OAA + H + NADH 3. 2PG <=> H2O + PEP 4. AKG + VAL <=> GLU + 3M-2-OB 5. AKG + NAD + CoA<=> SUC-CoA + CO2 + NADH | Transaminase (alaT) Malate dehydrogenase (mdh) Enolase (eno) Transaminase (ywaA) α-Ketoglutarate dehydrogenase (odhB) | 0.06 | 1.798 |
| Authors | Concentration (g/L) | Medium |
|---|---|---|
| Araujo et al., 2005 [10] | 0.17 | Potato-dextrose (PDA) |
| Zaidi et al., 2006 [11] | 0.055 | M9 + Glucose |
| Sivasakthi et al., 2013 [50] | 0.025 | Agar Nutriente |
| Alfonso et al., 2021 [30] | 0.086 | M9 + Propionate |
| This work | 0.31 | M9 + Propionate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillo-Alfonso, F.; Quintana-Menéndez, A.; Vigueras-Ramírez, G.; Sales-Cruz, A.M.; Rosales-Colunga, L.M.; Olivares-Hernández, R. Analysis of the Propionate Metabolism in Bacillus subtilis during 3-Indolacetic Production. Microorganisms 2022, 10, 2352. https://doi.org/10.3390/microorganisms10122352
Castillo-Alfonso F, Quintana-Menéndez A, Vigueras-Ramírez G, Sales-Cruz AM, Rosales-Colunga LM, Olivares-Hernández R. Analysis of the Propionate Metabolism in Bacillus subtilis during 3-Indolacetic Production. Microorganisms. 2022; 10(12):2352. https://doi.org/10.3390/microorganisms10122352
Chicago/Turabian StyleCastillo-Alfonso, Freddy, Alejandro Quintana-Menéndez, Gabriel Vigueras-Ramírez, Alfonso Mauricio Sales-Cruz, Luis Manuel Rosales-Colunga, and Roberto Olivares-Hernández. 2022. "Analysis of the Propionate Metabolism in Bacillus subtilis during 3-Indolacetic Production" Microorganisms 10, no. 12: 2352. https://doi.org/10.3390/microorganisms10122352