

Genome and Transcriptome Analysis to Elucidate the Biocontrol Mechanism of Bacillus amyloliquefaciens XJ5 against Alternaria solani
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. In Vitro Antifungal Activity Assays
2.3. Antifungal Activity Assessment of Crude Protein Extracts from XJ5
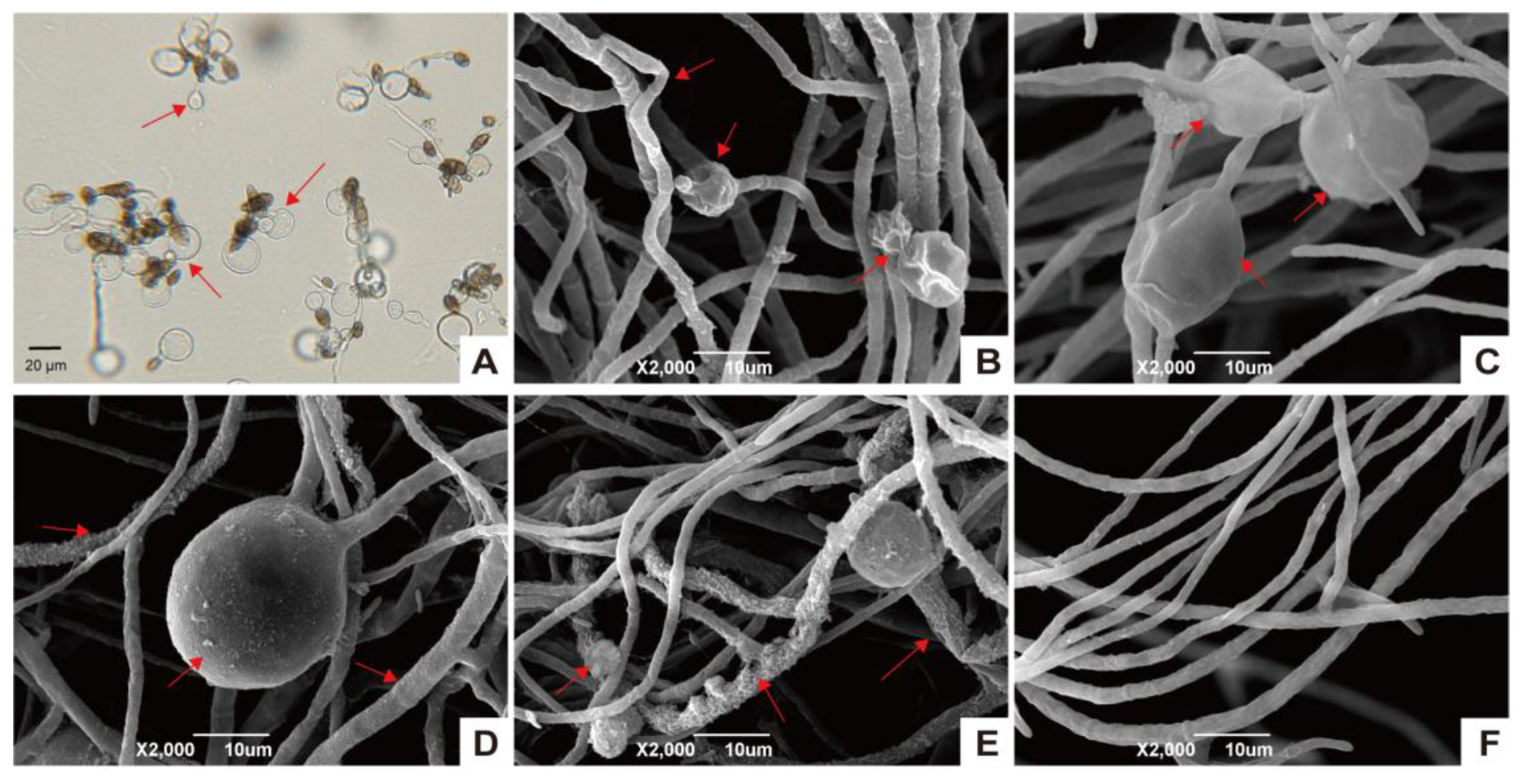
2.4. Light Microscopy and Scanning Electron Microscopy (SEM) of A. solani
2.5. DNA Extraction, Genome Sequencing, Assembly, and Annotation
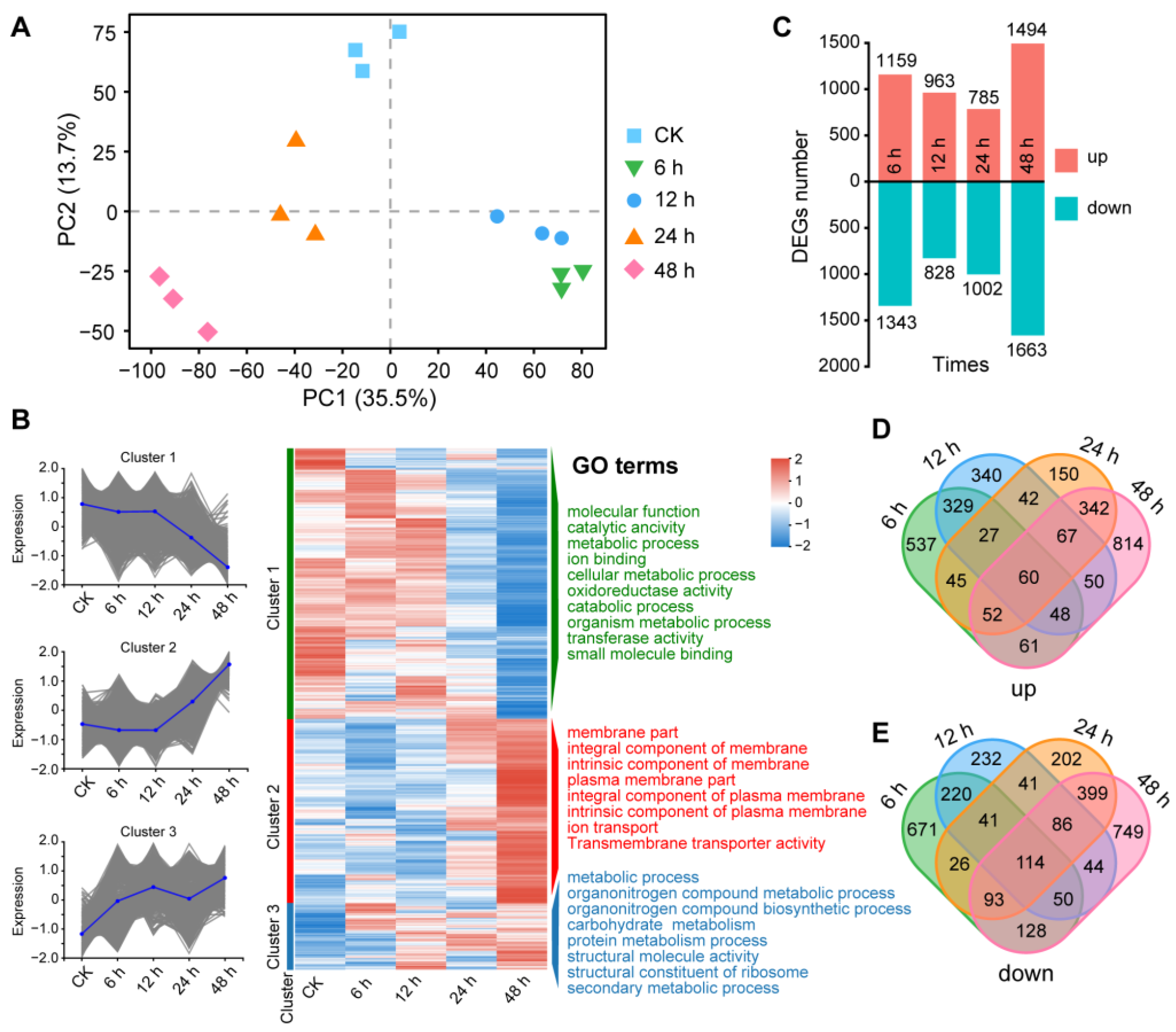
2.6. Total RNA Extraction, RNA Sequencing, and Transcriptomic Analysis
2.7. Real-Time Quantitative Reverse Transcription PCR (qRT-PCR) Analysis
3. Results
3.1. Morphological Observation and Antifungal Activity of B. amyloliquefaciens XJ5
3.2. XJ5 Inhibits Conidia Germination and Disturbs the Hyphal Structure of A. solani
3.3. Genome Features of B. amyloliquefaciens XJ5
3.4. Potential Functional Genes Involved in Biocontrol Traits
3.5. Transcriptomic Changes in A. solani Treated with B. amyloliquefaciens XJ5
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chaerani, R.; Voorrips, R.E. Tomato early blight (Alternaria solani): The pathogen, genetics, and breeding for resistance. J. Gen. Plant Pathol. 2006, 72, 335–347. [Google Scholar] [CrossRef]
- Lahkar, J.; Borah, S.N.; Deka, S.; Ahmed, G. Biosurfactant of Pseudomonas aeruginosa JS29 against Alternaria solani: The causal organism of early blight of tomato. BioControl 2015, 60, 401–411. [Google Scholar] [CrossRef]
- Gudmestad, N.C.; Arabiat, S.; Miller, J.S.; Pasche, J.S. Prevalence and impact of SDHI fungicide resistance in Alternaria solani. Plant Dis. 2013, 97, 952–960. [Google Scholar] [CrossRef] [Green Version]
- Leiminger, J.H.; Adolf, B.; Hausladen, H. Occurrence of the F 129L mutation in Alternaria solani populations in Germany in response to QoI application, and its effect on sensitivity. Plant Pathol. 2014, 63, 640–650. [Google Scholar] [CrossRef]
- Wharton, P.; Fairchild, K.; Belcher, A.; Wood, E. First report of in-vitro boscalid-resistant isolates of Alternaria solani causing early blight of potato in Idaho. Plant Dis. 2012, 96, 454. [Google Scholar] [CrossRef]
- Dang, J.; Gleason, M.L.; Wang, B.; Feng, J. Effects of Peganum harmala extracts and synthetic chemical fungicides on controlling early blight of tomato in the central shaanxi plain of China. Crop Prot. 2023, 166, 106177. [Google Scholar] [CrossRef]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Iqbal, S.; Begum, F.; Rabaan, A.A.; Aljeldah, M.; Al Shammari, B.R.; Alawfi, A.; Alshengeti, A.; Sulaiman, T.; Khan, A. Classification and multifaceted potential of secondary metabolites produced by Bacillus subtilis group: A comprehensive review. Molecules 2023, 28, 927. [Google Scholar]
- Dame, Z.T.; Rahman, M.; Islam, T. Bacilli as sources of agrobiotechnology: Recent advances and future directions. Green Chem. Lett. Rev. 2021, 14, 246–271. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Farzaneh, M.; Shi, Z.-Q.; Ahmadzadeh, M.; Hu, L.-B.; Ghassempour, A. Inhibition of the Aspergillus flavus growth and aflatoxin B1 contamination on pistachio nut by fengycin and surfactin-producing Bacillus subtilis UTBSP1. Plant Pathol. J. 2016, 32, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Li, B.; Zhang, N.; Waseem, R.; Shen, Q.; Huang, Q. Production of bacillomycin-and macrolactin-type antibiotics by Bacillus amyloliquefaciens NJN-6 for suppressing soilborne plant pathogens. J. Agric. Food Chem. 2012, 60, 2976–2981. [Google Scholar] [CrossRef] [PubMed]
- Béchet, M.; Caradec, T.; Hussein, W.; Abderrahmani, A.; Chollet, M.; Leclère, V.; Dubois, T.; Lereclus, D.; Pupin, M.; Jacques, P. Structure, biosynthesis, and properties of kurstakins, nonribosomal lipopeptides from Bacillus spp. Appl. Microbiol. Biotechnol. 2012, 95, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, Y.; Fu, X.; Li, Y.; Wang, Q. Isolation and characterization of Bacillus amyloliquefaciens PG12 for the biological control of apple ring rot. Postharvest Biol. Technol. 2016, 115, 113–121. [Google Scholar] [CrossRef]
- Wu, L.; Wu, H.; Chen, L.; Xie, S.; Zang, H.; Borriss, R.; Gao, X. Bacilysin from Bacillus amyloliquefaciens FZB42 has specific bactericidal activity against harmful algal bloom species. Appl. Environ. Microbiol. 2014, 80, 7512–7520. [Google Scholar] [CrossRef] [Green Version]
- Hammami, I.; Jaouadi, B.; Bacha, A.B.; Rebai, A.; Bejar, S.; Nesme, X.; Rhouma, A. Bacillus subtilis bacteriocin Bac 14B with a broad inhibitory spectrum: Purification, amino acid sequence analysis, and physicochemical characterization. Biotechnol. Bioprocess Eng. 2012, 17, 41–49. [Google Scholar] [CrossRef]
- Chen, X.-H.; Vater, J.; Piel, J.; Franke, P.; Scholz, R.; Schneider, K.; Koumoutsi, A.; Hitzeroth, G.; Grammel, N.; Strittmatter, A.W. Structural and functional characterization of three polyketide synthase gene clusters in Bacillus amyloliquefaciens FZB42. J. Bacteriol. 2006, 188, 4024–4036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Mutar, D.M.K.; Alzawar, N.S.A.; Noman, M.; Azizullah; Li, D.; Song, F. Suppression of Fusarium wilt in watermelon by Bacillus amyloliquefaciens DHA55 through extracellular production of antifungal lipopeptides. J. Fungi 2023, 9, 336. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-r.; Ling, X.-n.; Peng, S.-y.; Tan, M.-h.; Yan, L.-q.; Liang, Y.-y.; Li, G.-h.; Li, K.-t. A marine lipopeptides-producing Bacillus amyloliquefaciens HY2-1 with a broad-spectrum antifungal and antibacterial activity and its fermentation kinetics study. World J. Microbiol. Biotechnol. 2023, 39, 196. [Google Scholar] [CrossRef] [PubMed]
- Leelasuphakul, W.; Sivanunsakul, P.; Phongpaichit, S. Purification, characterization and synergistic activity of β-1, 3-glucanase and antibiotic extract from an antagonistic Bacillus subtilis NSRS 89-24 against rice blast and sheath blight. Enzyme Microb. Technol. 2006, 38, 990–997. [Google Scholar] [CrossRef]
- Khochamit, N.; Siripornadulsil, S.; Sukon, P.; Siripornadulsil, W. Antibacterial activity and genotypic–phenotypic characteristics of bacteriocin-producing Bacillus subtilis KKU213: Potential as a probiotic strain. Microbiol. Res. 2015, 170, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Chakdar, H.; Pandiyan, K.; Thapa, S.; Shahid, M.; Singh, A.; Srivastava, A.K.; Saxena, A.K. Bacterial chitinases: Genetics, engineering and applications. World J. Microbiol. Biotechnol. 2022, 38, 252. [Google Scholar] [CrossRef] [PubMed]
- Essghaier, B.; Zouaoui, M.; Najjari, A.; Sadfi, N. Potentialities and characterization of an antifungal chitinase produced by a halotolerant Bacillus licheniformis. Curr. Microbiol. 2021, 78, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.S.; Rehman, A.; Khan, I.U.; Khan, T.A.; Jamil, M.; Rha, E.S.; Anees, M. Thermo-neutrophilic cellulases and chitinases characterized from a novel putative antifungal biocontrol agent: Bacillus subtilis TD11. PLoS ONE 2023, 18, e0281102. [Google Scholar] [CrossRef]
- Lim, S.M.; Yoon, M.-Y.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Shin, T.S.; Park, H.W.; Yu, N.H.; Kim, Y.H.; Kim, J.-C. Diffusible and volatile antifungal compounds produced by an antagonistic Bacillus velezensis G341 against various phytopathogenic fungi. Plant Pathol. J. 2017, 33, 488. [Google Scholar] [CrossRef] [Green Version]
- Poulaki, E.G.; Tjamos, S.E. Bacillus species: Factories of plant protective volatile organic compounds. J. Appl. Bacteriol. 2023, 134, lxad037. [Google Scholar] [CrossRef]
- Tian, Y.; Ji, S.; Zhang, E.; Chen, Y.; Xu, G.; Chen, X.; Fan, J.; Tang, X. Complete genome analysis of Bacillus subtilis TY-1 reveals its biocontrol potential against tobacco bacterial wilt. Mar. Genom. 2023, 68, 101018. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Bowman, M.J.; Schisler, D.A. Genomic analysis and secondary metabolite production in Bacillus amyloliquefaciens AS 43.3: A biocontrol antagonist of Fusarium head blight. Biol. Control 2013, 64, 166–175. [Google Scholar]
- Douriet-Gámez, N.R.; Maldonado-Mendoza, I.E.; Ibarra-Laclette, E.; Blom, J.; Calderón-Vázquez, C.L. Genomic analysis of Bacillus sp. strain B25, a biocontrol agent of maize pathogen Fusarium verticillioides. Curr. Microbiol. 2018, 75, 247–255. [Google Scholar] [CrossRef]
- Zhao, Y.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Folly, Y.M.E. Antagonistic action of Bacillus subtilis strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar]
- Liu, D.; Li, K.; Hu, J.; Wang, W.; Liu, X.; Gao, Z. Biocontrol and action mechanism of Bacillus amyloliquefaciens and Bacillus subtilis in soybean phytophthora blight. Int. J. Mol. Sci. 2019, 20, 2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, D.; Cai, X.; Yang, C.; Xie, L.; Qin, G.; Zhang, M.; Huang, Y.; Gong, G.; Chang, X.; Chen, H. Studies on the control effect of Bacillus subtilis on wheat powdery mildew. Pest Manag. Sci. 2021, 77, 4375–4382. [Google Scholar]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O. Comparative analysis of the complete genome sequence of the plant growth–promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- Rückert, C.; Blom, J.; Chen, X.; Reva, O.; Borriss, R. Genome sequence of B. amyloliquefaciens type strain DSM7T reveals differences to plant-associated B. amyloliquefaciens FZB42. J. Bacteriol. 2011, 155, 78–85. [Google Scholar]
- Wang, D.; Luo, W.-Z.; Zhang, D.-D.; Li, R.; Kong, Z.-Q.; Song, J.; Dai, X.-F.; Alkan, N.; Chen, J.-Y. Insights into the biocontrol function of a Burkholderia gladioli strain against Botrytis cinerea. Microbiol. Spectrum 2023, 11, e04805-22. [Google Scholar]
- Ren, J.; He, W.; Li, C.; He, S.; Niu, D. Purification and identification of a novel antifungal protein from Bacillus subtilis XB-1. World J. Microbiol. Biotechnol. 2019, 35, 150. [Google Scholar] [CrossRef]
- Opabode, J.T.; Raji, I.B. Evaluation of genomic DNA-extraction methods for molecular analysis of solanecio biafrae. Int. J. Veg. Sci. 2019, 25, 111–123. [Google Scholar] [CrossRef]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA genes in genomic sequences. In Gene Prediction: Methods and Protocols; Kollmar, M., Ed.; Springer: New York, NY, USA, 2019; pp. 1–14. [Google Scholar]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Cui, X.; Lu, Z.; Wang, S.; Wang, J.; Gao, X. CMsearch: Simultaneous exploration of protein sequence space and structure space improves not only protein homology detection but also protein structure prediction. Bioinformatics 2016, 32, i332–i340. [Google Scholar] [CrossRef] [Green Version]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Rajarammohan, S. Transcriptome Analysis of the Necrotrophic Pathogen Alternaria brassicae Reveals Insights into Its Pathogenesis in Brassica juncea. Microbiol. Spectr. 2023, 11, e02939-22. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [PubMed] [Green Version]
- Puertolas, A.; Bonants, P.J.M.; Boa, E.; Woodward, S. Application of real-time PCR for the detection and quantification of oomycetes in ornamental nursery stock. J. Fungi 2021, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Galileya Medison, R.; Zheng, T.-w.; Meng, X.-j.; Sun, Z.-x.; Zhou, Y. Biocontrol potential of Bacillus amyloliquefaciens YZU-SG146 from Fraxinus hupehensis against Verticillium wilt of cotton. Biol. Control 2023, 183, 105246. [Google Scholar] [CrossRef]
- Marković, S.; Milovanović, T.P.; Jelušić, A.; Iličić, R.; Medić, O.; Berić, T.; Stanković, S. Biological control of major pathogenic bacteria of potato by Bacillus amyloliquefaciens strains SS-12.6 and SS-38.4. Biol. Control 2023, 182, 105238. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J.; Liu, T.; Li, X.; Gao, J.; Jiang, Y.; Chen, C.-Q. Bacillus amyloliquefaciens FG14 as a potential biocontrol strain against rusty root rot of Panax ginseng, and its impact on the rhizosphere microbial community. Biol. Control 2023, 182, 105221. [Google Scholar] [CrossRef]
- Hanif, A.; Zhang, F.; Li, P.; Li, C.; Xu, Y.; Zubair, M.; Zhang, M.; Jia, D.; Zhao, X.; Liang, J. Fengycin produced by Bacillus amyloliquefaciens FZB42 inhibits Fusarium graminearum growth and mycotoxins biosynthesis. Toxins 2019, 11, 295. [Google Scholar] [CrossRef] [Green Version]
- Zouari, I.; Jlaiel, L.; Tounsi, S.; Trigui, M. Biocontrol activity of the endophytic Bacillus amyloliquefaciens strain CEIZ-11 against Pythium aphanidermatum and purification of its bioactive compounds. Biol. Control 2016, 100, 54–62. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, S.; Wang, D.; Xue, J.; Guo, D.; Song, X.; Zhang, F.; Huang, S.; Luan, C. Complete genome sequence of Bacillus amyloliquefaciens B15 isolated from grape skin, a strain of strong inhibitory activity against fungi. J. Bacteriol. 2016, 228, 28–29. [Google Scholar]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shao, J.; Li, B.; Yan, X.; Shen, Q.; Zhang, R. Contribution of bacillomycin D in Bacillus amyloliquefaciens SQR9 to antifungal activity and biofilm formation. Appl. Environ. Microbiol. 2013, 79, 808–815. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Li, Q.; Xu, Z.; Zhang, N.; Shen, Q.; Zhang, R. Responses of beneficial Bacillus amyloliquefaciens SQR9 to different soilborne fungal pathogens through the alteration of antifungal compounds production. Front. Microbiol. 2014, 5, 636. [Google Scholar] [CrossRef] [Green Version]
- Cantu, D.; Carl Greve, L.; Labavitch, J.M.; Powell, A.L.T. Characterization of the cell wall of the ubiquitous plant pathogen Botrytis cinerea. Mycol. Res. 2009, 113, 1396–1403. [Google Scholar] [CrossRef]
- Swiontek Brzezinska, M.; Kalwasińska, A.; Świątczak, J.; Żero, K.; Jankiewicz, U. Exploring the properties of chitinolytic Bacillus isolates for the pathogens biological control. Microb. Pathog. 2020, 148, 104462. [Google Scholar] [CrossRef]
- Gong, Q.; Zhang, C.; Lu, F.; Zhao, H.; Bie, X.; Lu, Z. Identification of bacillomycin D from Bacillus subtilis fmbJ and its inhibition effects against Aspergillus flavus. Food Control 2014, 36, 8–14. [Google Scholar] [CrossRef]
- Duan, Y.; Chen, R.; Zhang, R.; Jiang, W.; Chen, X.; Yin, C.; Mao, Z. Isolation, identification, and antibacterial mechanisms of Bacillus amyloliquefaciens QSB-6 and its effect on plant roots. Front. Microbiol. 2021, 12, 746799. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fang, W.; Zhang, L.; Ouyang, H.; Zhou, H.; Luo, Y.; Jin, C. Class IIC α-mannosidase AfAms1 is required for morphogenesis and cellular function in Aspergillus fumigatus. Glycobiology 2009, 19, 624–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhou, H.; Ouyang, H.; Li, Y.; Jin, C. Afcwh41 is required for cell wall synthesis, conidiation, and polarity in Aspergillus fumigatus. FEMS Microbiol. Lett. 2008, 289, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, A.; Fernando, L.D.; Fang, W.; Dickwella Widanage, M.C.; Wei, P.; Jin, C.; Fontaine, T.; Latge, J.P.; Wang, T. A molecular vision of fungal cell wall organization by functional genomics and solid-state NMR. Nat. Commun. 2021, 12, 6346. [Google Scholar] [CrossRef]
- Zhou, H.; Hu, H.; Zhang, L.; Li, R.; Ouyang, H.; Ming, J.; Jin, C. O-Mannosyltransferase 1 in Aspergillus fumigatus (AfPmt1p) is crucial for cell wall integrity and conidium morphology, especially at an elevated temperature. Eukaryot. Cell 2008, 6, 2260–2268. [Google Scholar] [CrossRef] [Green Version]
- Veal, E.A.; Toone, W.M.; Jones, N.; Morgan, B.A. Distinct roles for glutathione S-transferases in the oxidative stress response in Schizosaccharomyces pombe. J. Biol. Chem. 2002, 277, 35523–35531. [Google Scholar] [CrossRef] [Green Version]
- Garcerá, A.; Barreto, L.; Piedrafita, L.; Tamarit, J.; Herrero, E. Saccharomyces cerevisiae cells have three Omega class glutathione S-transferases acting as 1-Cys thiol transferases. Biochem. J. 2006, 398, 187–196. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster ID | Type | Similar Cluster | Similarity % | Number of Genes |
|---|---|---|---|---|
| Cluster1 | NRPS | Locillomycin | 28 | 46 |
| Cluster2 | NRPS | Surfactin | 91 | 43 |
| Cluster3 | PKS-like | Butirosin A/ButirosinB | 7 | 39 |
| Cluster 4 | Terpene | - | 23 | |
| Cluster5 | TransAT-PKS | Macrolactin H | 100 | 43 |
| Cluster6 | TransAT-PKS | Bacillaene | 100 | 44 |
| Cluster7 | NRPS | Fengycin | 100 | 67 |
| Cluster8 | Terpene | - | - | 21 |
| Cluster9 | T3PKS | - | - | 62 |
| Cluster10 | TransAT-PKS | Difficidin | 100 | 41 |
| Cluster11 | NRPS | Bacillibactin | 100 | 48 |
| Cluster12 | NRPS | - | - | 38 |
| Cluster13 | Other | Bacilysin | 100 | 43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, F.; Chen, X.; Fu, Z.; Wang, X.; Guo, J.; Zhao, X.; Zhang, B. Genome and Transcriptome Analysis to Elucidate the Biocontrol Mechanism of Bacillus amyloliquefaciens XJ5 against Alternaria solani. Microorganisms 2023, 11, 2055. https://doi.org/10.3390/microorganisms11082055
Mu F, Chen X, Fu Z, Wang X, Guo J, Zhao X, Zhang B. Genome and Transcriptome Analysis to Elucidate the Biocontrol Mechanism of Bacillus amyloliquefaciens XJ5 against Alternaria solani. Microorganisms. 2023; 11(8):2055. https://doi.org/10.3390/microorganisms11082055
Chicago/Turabian StyleMu, Fan, Xu Chen, Zhenxin Fu, Xue Wang, Jiexin Guo, Xiaojun Zhao, and Baojun Zhang. 2023. "Genome and Transcriptome Analysis to Elucidate the Biocontrol Mechanism of Bacillus amyloliquefaciens XJ5 against Alternaria solani" Microorganisms 11, no. 8: 2055. https://doi.org/10.3390/microorganisms11082055