
Carotenoid Cocktail Produced by An Antarctic Soil Flavobacterium with Biotechnological Potential

 , ,
, ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
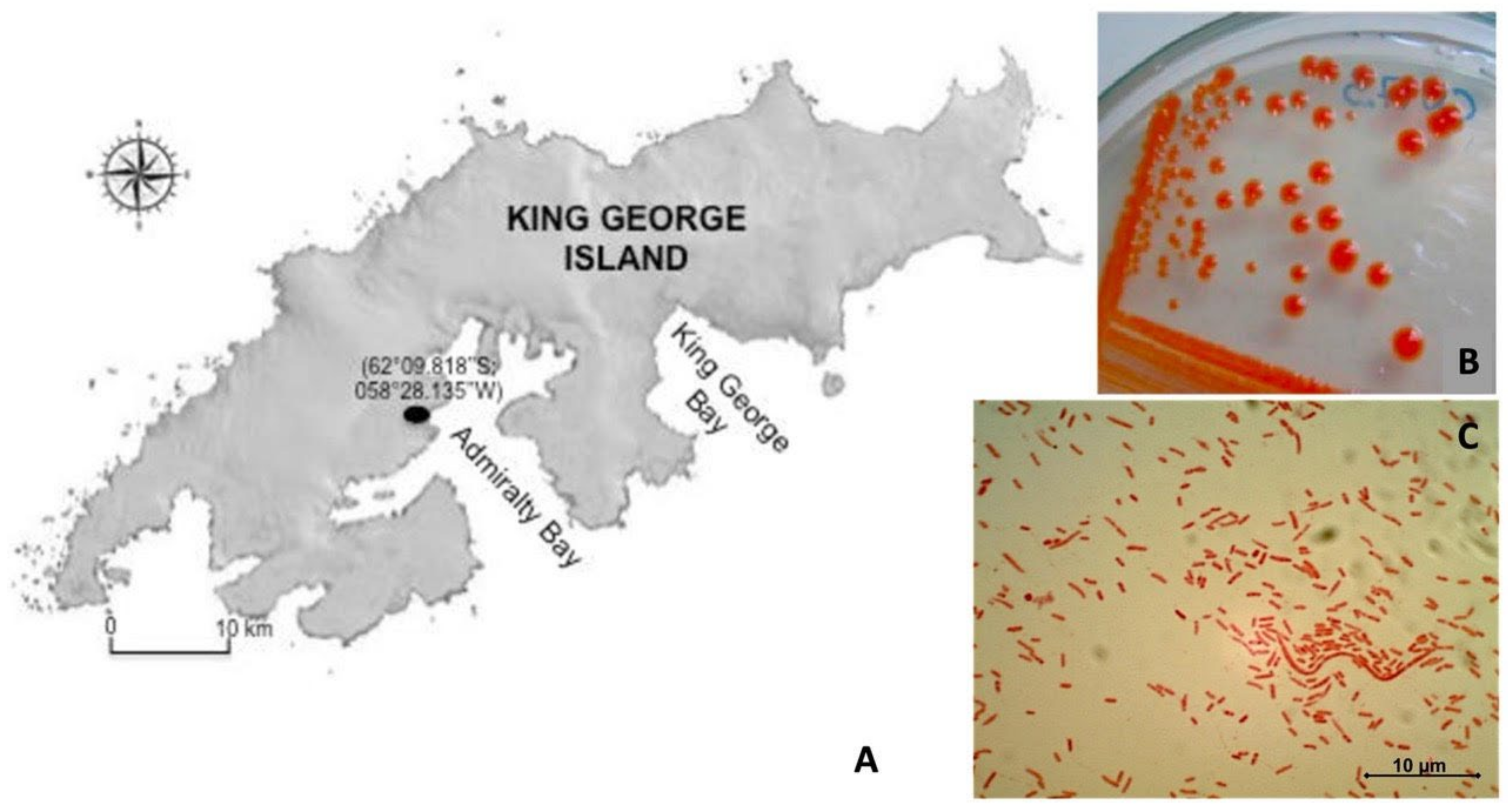
2.1. Bacterial Sample and Morphological Characterization
2.2. DNA Extraction and Amplification of the 16S rRNA Gene of CN7 Strain
2.3. CN7 Pigment Extraction
2.4. High-Performance Liquid Chromatography (HPLC) and UV-Vis Detection
2.5. Liquid Chromatography–UltraViolet-Mass Spectrometry (LC–UV-MS/MS) Analysis for Pigment Identification
3. Results
3.1. Characteristics of the Isolated CN7 Strain
3.2. CN7 Strain Identification
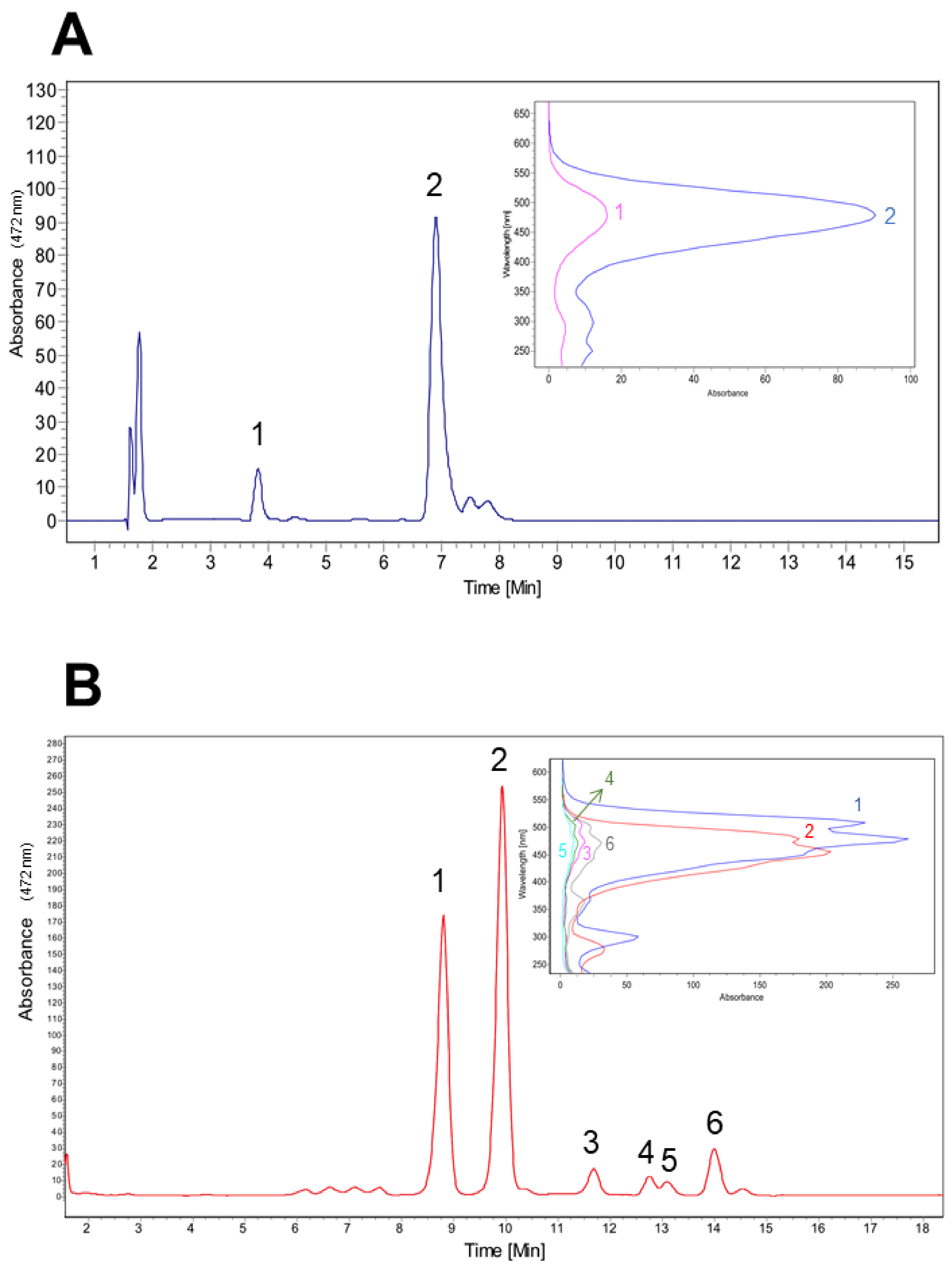
3.3. Carotenoids Fractionation from CN7 Strain
3.4. Carotenoids Identification from Antarctic Isolated CN7
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Margesin, R.; Miteva, V. Diversity and ecology of psychrophilic microorganisms. Res. Microbiol. 2011, 162, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Dieser, M.; Greenwood, M.; Foreman, C.M. Carotenoid pigmentation in Antarctic heterotrophic bacteria as a strategy to withstand environmental stresses. Arctic Antarct. Alp. Res. 2010, 42, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Reis-Mansur, M.C.P.P.; Cardoso-Rurr, J.S.; Silva, J.V.M.A.; de Souza, G.R.; da Silva Cardoso, V.; Mansoldo, F.R.P.; Pinheiro, Y.; Schultz, J.; Lopez Balottin, L.B.; da Silva, A.J.R.; et al. Carotenoids from UV-resistant Antarctic Microbacterium sp. LEMMJ01. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.; Gomes, N.C.M.; Henriques, I.; Almeida, A.; Correia, A.; Cunha, Â. Contribution of reactive oxygen species to UV-B-induced damage in bacteria. J. Photochem. Photobiol. B Biol. 2012, 117, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.; Margesin, R. Psychrophilic lifestyles: Mechanisms of adaptation and biotechnological tools. Appl. Microbiol. Biotechnol. 2019, 103, 2857–2871. [Google Scholar] [CrossRef] [PubMed]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.-F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Martín-Cerezo, M.L.; García-López, E.; Cid, C. Isolation and Identification of a Red Pigment from the Antarctic Bacterium Shewanella frigidimarina. Protein Pept. Lett. 2015, 22, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, Y.; Wang, N.; Jiao, N.; Xu, B.; Liu, X. Variation with depth of the abundance, diversity and pigmentation of culturable bacteria in a deep ice core from the Yuzhufeng Glacier, Tibetan Plateau. Extremophiles 2018, 22, 29–38. [Google Scholar] [CrossRef]
- Perez-Fons, L.; Steiger, S.; Khaneja, R.; Bramley, P.M.; Cutting, S.M.; Sandmann, G.; Fraser, P.D. Identification and the developmental formation of carotenoid pigments in the yellow/orange Bacillus spore-formers. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 177–185. [Google Scholar] [CrossRef]
- Ramesh, C.; Vinithkumar, N.; Kirubagaran, R. Marine pigmented bacteria: A prospective source of antibacterial compounds. J. Nat. Sci. Biol. Med. 2019, 10, 104. [Google Scholar] [CrossRef]
- Varela, J.C.; Pereira, H.; Vila, M.; León, R. Production of carotenoids by microalgae: Achievements and challenges. Photosynth. Res. 2015, 125, 423–436. [Google Scholar] [CrossRef]
- de Carvalho, C.C.C.R.; Caramujo, M.J. Carotenoids in aquatic ecosystems and aquaculture: A colorful business with implications for human health. Front. Mar. Sci. 2017, 4, 93. [Google Scholar] [CrossRef] [Green Version]
- Venil, C.K.; Zakaria, Z.A.; Ahmad, W.A. Bacterial pigments and their applications. Process Biochem. 2013, 48, 1065–1079. [Google Scholar] [CrossRef]
- Khalid, M.; Saeed-ur-Rahman; Iqbal, H.M.N.; Huang, D. Biosynthesis and biomedical perspectives of carotenoids with special reference to human health-related applications. Biocatal. Agric. Biotechnol. 2019, 17, 399–407. [Google Scholar] [CrossRef]
- Sefc, K.M.; Brown, A.C.; Clotfelter, E.D. Carotenoid-based coloration in cichlid fishes. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 173, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Maan, M.E.; Sefc, K.M. Colour variation in cichlid fish: Developmental mechanisms, selective pressures and evolutionary consequences. Semin. Cell Dev. Biol. 2013, 24, 516–528. [Google Scholar] [CrossRef] [Green Version]
- Arous, W.H.; El-bermawi, N.M.; Shaltout, O.E.; Essa, M.A.E. Effect of Adding Different Carotenoid Sources on Growth Performance, Pigmentation, Stress Response and Quality in Red Tilapia (Oreochromis Spp.). Middle East J. Appl. Sci. 2014, 4, 988–999. [Google Scholar]
- Torrissen, O.J.; Christiansen, R. Requirements for carotenoids in fish diets. J. Appl. Ichthyol. 1995, 11, 225–230. [Google Scholar] [CrossRef]
- Shahidi, F.; Brown, J.A. Carotenoid Pigments in Seafoods and Aquaculture. Crit. Rev. Food Sci. Nutr. 1998, 38, 1–67. [Google Scholar] [CrossRef]
- Beveridge, T. Use of the Gram stain in microbiology. Biotech. Histochem. 2001, 76, 111–118. [Google Scholar] [CrossRef]
- Sambrook, J.F.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; ISBN 9780879695774. [Google Scholar]
- Hauschild, T.; Stepanovic, S. Identification of Staphylococcus spp. by PCR-Restriction Fragment Length Polymorphism Analysis of dnaJ Gene. J. Clin. Microbiol. 2008, 46, 3875–3879. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.K.; Chung, E.J.; Kim, J.-C.; Choi, G.J.; Jang, K.S.; Chung, Y.R.; Cho, K.Y.; Lee, S.-W. Characterization of a Forest Soil Metagenome Clone That Confers Indirubin and Indigo Production on Escherichia coli. Appl. Environ. Microbiol. 2005, 71, 7768–7777. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Chenna, R. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- De Rosso, V.V.; Mercadante, A.Z. Identification and quantification of carotenoids, by HPLC-PDA-MS/MS, from Amazonian fruits. J. Agric. Food Chem. 2007, 55, 5062–5072. [Google Scholar] [CrossRef]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Zaghdoudi, K.; Ngomo, O.; Vanderesse, R.; Arnoux, P.; Myrzakhmetov, B.; Frochot, C.; Guiavarc’h, Y. Extraction, Identification and Photo-Physical Characterization of Persimmon (Diospyros kaki L.) Carotenoids. Foods 2017, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweiggert, U.; Kammerer, D.R.; Carle, R.; Schieber, A. Characterization of carotenoids and carotenoid esters in red pepper pods (Capsicum annuum L.) by high-performance liquid chromatography/atmospheric pressure chemical ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 2617–2628. [Google Scholar] [CrossRef] [PubMed]
- Crupi, P.; Milella, R.A.; Antonacci, D. Simultaneous HPLC-DAD-MS (ESI+) determination of structural and geometrical isomers of carotenoids in mature grapes. J. Mass Spectrom. 2010, 45, 971–980. [Google Scholar] [CrossRef]
- Kosourov, S.; Murukesan, G.; Jokela, J.; Allahverdiyeva, Y. Carotenoid biosynthesis in Calothrix sp. 336/3: Composition of carotenoids on full medium, during diazotrophic growth and after long-term H2 photoproduction. Plant Cell Physiol. 2016, 57, 2269–2282. [Google Scholar] [CrossRef] [Green Version]
- de Azevedo-Meleiro, C.H.; Rodriguez-Amaya, D.B. Qualitative and quantitative differences in the carotenoid composition of yellow and red peppers determined by HPLC-DAD-MS. J. Sep. Sci. 2009, 32, 3652–3658. [Google Scholar] [CrossRef]
- van Breemen, R.B.; Dong, L.; Pajkovic, N.D. Atmospheric pressure chemical ionization tandem mass spectrometry of carotenoids. Int. J. Mass Spectrom. 2012, 312, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Mi, J.; Jia, K.-P.; Wang, J.Y.; Al-Babili, S. A rapid LC–MS method for qualitative and quantitative profiling of plant apocarotenoids. Anal. Chim. Acta 2018, 1035, 87–95. [Google Scholar] [CrossRef]
- Roesler, C.S.; Culbertson, C.W.; Etheridge, S.M.; Goericke, R.; Kiene, R.P.; Miller, L.G.; Oremland, R.S. Distribution, production, and ecophysiology of Picocystis strain ML in Mono Lake, California. Limnol. Oceanogr. 2002, 47, 440–452. [Google Scholar] [CrossRef]
- de Faria, A.F.; de Rosso, V.V.; Mercadante, A.Z. Carotenoid Composition of Jackfruit (Artocarpus heterophyllus), Determined by HPLC-PDA-MS/MS. Plant Foods Hum. Nutr. 2009, 64, 108–115. [Google Scholar] [CrossRef]
- Ferreres, F.; Pereira, D.M.; Gil-Izquierdo, A.; Valentão, P.; Botelho, J.; Mouga, T.; Andrade, P.B. HPLC-PAD-atmospheric pressure chemical ionization-MS metabolite profiling of cytotoxic carotenoids from the echinoderm Marthasterias glacialis (spiny sea-star). J. Sep. Sci. 2010, 33, 2250–2257. [Google Scholar] [CrossRef]
- Gentili, A.; Caretti, F.; Bellante, S.; Ventura, S.; Canepari, S.; Curini, R. Comprehensive Profiling of Carotenoids and Fat-Soluble Vitamins in Milk from Different Animal Species by LC-DAD-MS/MS Hyphenation. J. Agric. Food Chem. 2013, 61, 1628–1639. [Google Scholar] [CrossRef]
- Bóna-Lovász, J.; Bóna, A.; Ederer, M.; Sawodny, O.; Ghosh, R. A Rapid Method for the Extraction and Analysis of Carotenoids and Other Hydrophobic Substances Suitable for Systems Biology Studies with Photosynthetic Bacteria. Metabolites 2013, 3, 912–930. [Google Scholar] [CrossRef]
- Campos Chisté, R.; Zerlotti Mercadante, A. Identification and quantification, by HPLC-DAD-MS/MS, of carotenoids and phenolic compounds from the Amazonian fruit Caryocar villosum. J. Agric. Food Chem. 2012, 60, 5884–5892. [Google Scholar] [CrossRef]
- Aberoumand, A. A review article on edible pigments properties and sources as natural biocolorants in foodstuff and food industry. J. Dairy Food Sci. 2011, 6, 71–78. [Google Scholar]
- Kirti, K.; Amita, S.; Priti, S.; Mukesh Kumar, A.; Jyoti, S. Colorful World of Microbes: Carotenoids and Their Applications. Adv. Biol. 2014, 2014, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Vila, E.; Hornero-Méndez, D.; Azziz, G.; Lareo, C.; Saravia, V. Carotenoids from heterotrophic bacteria isolated from Fildes Peninsula, King George Island, Antarctica. Biotechnol. Reports 2019, 21, e00306. [Google Scholar] [CrossRef] [PubMed]
- Vila, E.; Hornero-Méndez, D.; Lareo, C.; Saravia, V. Biotechnological production of zeaxanthin by an Antarctic Flavobacterium: Evaluation of culture conditions. J. Biotechnol. 2020, 319, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, P.; Larson, A.J.; Bernstein, P.S. Factorial analysis of tricarboxylic acid cycle intermediates for optimization of zeaxanthin production from Flavobacterium multivorum. J. Appl. Microbiol. 2004, 96, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Chun, J. Flavobacterium weaverense sp. nov. and Flavobacterium segetis sp. nov., novel psychrophiles isolated from the Antarctic. Int. J. Syst. Evol. Microbiol. 2006, 56, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [Green Version]
- Ram, S.; Mitra, M.; Shah, F.; Tirkey, S.R.; Mishra, S. Bacteria as an alternate biofactory for carotenoid production: A review of its applications, opportunities and challenges. J. Funct. Foods 2020, 67, 103867. [Google Scholar] [CrossRef]
- Gierhart, D.L. Production of Zeaxanthin and Zeaxanthin-Containing; Applied Food Biotechnology, Inc.: Fenton, MO, USA, 1994. [Google Scholar]
- Turcsi, E.; Nagy, V.; Deli, J. Study on the elution order of carotenoids on endcapped C18 and C30 reverse silica stationary phases. A review of the database. J. Food Compos. Anal. 2016, 47, 101–112. [Google Scholar] [CrossRef]
- Juturu, V.; Bowman, P.J.; Stringham, T.N.; Stringham, M.J. Bioavailability of lutein/zeaxanthin isomers and macular pigment optical density response to macular carotenoid supplementation: A randomized double blind placebo controlled study. New Front. Ophthalmol. 2016, 2, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Manikandan, R.; Thiagarajan, R.; Goutham, G.; Arumugam, M.; Beulaja, M.; Rastrelli, L.; Skalicka-Woźniak, K.; Habtemariam, S.; Orhan, I.E.; Nabavi, S.F.; et al. Zeaxanthin and ocular health, from bench to bedside. Fitoterapia 2016, 109, 58–66. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, J.H.; Lee, P.C. Flavobacterium kingsejongi sp. nov., a carotenoid-producing species isolated from Antarctic penguin faeces. Int. J. Syst. Evol. Microbiol. 2018, 68, 911–916. [Google Scholar] [CrossRef]
- Khan, I.; Debnath, S.C.; Yan, C.; Chen, C.; Xu, Y.; Wang, W.-J.; Yu, Y.-C.; Zheng, D.-Q.; Xu, J.-Z.; Wang, P.-M. Flavobacterium ajazii sp. nov., Isolated from Seaweed of Gouqi Island, China. Curr. Microbiol. 2020, 77, 2925–2932. [Google Scholar] [CrossRef]
- Liu, Q.; Li, W.; Liu, D.; Li, L.; Li, J.; Lv, N.; Liu, F.; Zhu, B.; Zhou, Y.; Xin, Y.; et al. Light stimulates anoxic and oligotrophic growth of glacial Flavobacterium strains that produce zeaxanthin. ISME J. 2021, 15, 1844–1857. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Z.; Sun, J.; Xue, C.; Mao, X. Biotechnological production of zeaxanthin by microorganisms. Trends Food Sci. Technol. 2018, 71, 225–234. [Google Scholar] [CrossRef]
- Ron, E.Y.C.; Plaza, M.; Kristjansdottir, T.; Sardari, R.R.R.; Bjornsdottir, S.H.; Gudmundsson, S.; Hreggvidsson, G.O.; Turner, C.; van Niel, E.W.J.; Nordberg-Karlsson, E. Characterization of carotenoids in Rhodothermus marinus. Microbiologyopen 2018, 7, e00536. [Google Scholar] [CrossRef] [Green Version]
- Kopsell, D.A.; Kopsell, D.E. Accumulation and bioavailability of dietary carotenoids in vegetable crops. Trends Plant Sci. 2006, 11, 499–507. [Google Scholar] [CrossRef]
- Chi, S.C.; Mothersole, D.J.; Dilbeck, P.; Niedzwiedzki, D.M.; Zhang, H.; Qian, P.; Vasilev, C.; Grayson, K.J.; Jackson, P.J.; Martin, E.C.; et al. Assembly of functional photosystem complexes in Rhodobacter sphaeroides incorporating carotenoids from the spirilloxanthin pathway. Biochim. Biophys. Acta Bioenerg. 2015, 1847, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Amaya, D.B. OMNI (Project): A Guide to Carotenoid Analysis in Foods; ILSI Press: Washington, DC, USA, 2001; ISBN 9781578810727. [Google Scholar]
- Schieber, A.; Weber, F. Handbook on Natural Pigments in Food and Beverages: Industrial Applications for Improving Food Color; Woodhead Publishing: Sawston, UK, 2016; ISBN 9780081003923. [Google Scholar]
- Matsumo, T.; Nagata, S.; Iwahashi, M.; Koikb, T.; Okada, M. Intensification of color of fancy red carp with zeaxanthin and myxoxanthophyll, major carotenoid constituents of spirulina. Nippon Suisan Gakkaishi 1979, 45, 627–632. [Google Scholar] [CrossRef]
- Alcaíno, J.; Baeza, M.; Cifuentes, V. Carotenoid Distribution in Nature. Subcell. Biochem. 2016, 79, 3–33. [Google Scholar] [CrossRef]
- Correa-Llantén, D.N.; Amenábar, M.J.; Blamey, J.M. Antioxidant capacity of novel pigments from an Antarctic bacterium. J. Microbiol. 2012, 50, 374–379. [Google Scholar] [CrossRef]
- Johnson, E.J. Role of lutein and zeaxanthin in visual and cognitive function throughout the lifespan. Nutr. Rev. 2014, 72, 605–612. [Google Scholar] [CrossRef]
- Moreno, J.A.; Díaz-Gómez, J.; Nogareda, C.; Angulo, E.; Sandmann, G.; Portero-Otin, M.; Serrano, J.C.E.; Twyman, R.M.; Capell, T.; Zhu, C.; et al. The distribution of carotenoids in hens fed on biofortified maize is influenced by feed composition, absorption, resource allocation and storage. Sci. Rep. 2016, 6, 35346. [Google Scholar] [CrossRef] [Green Version]
- Breitenbach, J.; Nogueira, M.; Farré, G.; Zhu, C.; Capell, T.; Christou, P.; Fleck, G.; Focken, U.; Fraser, P.D.; Sandmann, G. Engineered maize as a source of astaxanthin: Processing and application as fish feed. Transgenic Res. 2016, 25, 785–793. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, M.; Enfissi, E.M.A.; Martínez Valenzuela, M.E.; Menard, G.N.; Driller, R.L.; Eastmond, P.J.; Schuch, W.; Sandmann, G.; Fraser, P.D. Engineering of tomato for the sustainable production of ketocarotenoids and its evaluation in aquaculture feed. Proc. Natl. Acad. Sci. USA 2017, 114, 10876–10881. [Google Scholar] [CrossRef] [Green Version]
- Nadarajah, S.; Flaaten, O. Global aquaculture growth and institutional quality. Mar. Policy 2017, 84, 142–151. [Google Scholar] [CrossRef] [Green Version]
- García-Chavarría, M.; Lara-Flores, M. The use of carotenoid in aquaculture. Res. J. Fish. Hydrobiol. 2013, 8, 38–49. [Google Scholar]
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from marine organisms: Biological functions and industrial applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, N.R.; Fineran, P.C.; Leeper, F.J.; Salmond, G.P.C. The biosynthesis and regulation of bacterial prodiginines. Nat. Rev. Microbiol. 2006, 4, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Rehm, B.H.A. Bacterial polymers: Biosynthesis, modifications and applications. Nat. Rev. Microbiol. 2010, 8, 578–592. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | RT (min) | Area (%) | Precursor (m/z) | Fragments MS2 (m/z) | Identification | References | |||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 15.7 | 0.4 | 585.2 | 492.3 | 566.4 | 525.4 | 485.6 | Capsanthin | [31] |
| Lutein-5,6-epoxide | [32] | ||||||||
| Caloxanthin (=Hydroxy-zeaxanthin) | [33] | ||||||||
| 2 | 17.2 | 0.7 | 584.4 | 492.2 | 564.4 | 567.0 | 477.2 | Capsanthin | [34,35] |
| Lutein-5,6-epoxide | [32] | ||||||||
| Caloxanthin (=Hydroxy-zeaxanthin) | [33] | ||||||||
| 413.7 | 301.0 | 296.3 | 188.5 | 395.2 | Not identified | ||||
| 3 | 19.7 | 0.5 | 284.6 | 240.0 | 101.5 | 87.5 | 115.5 | 15-apo-carotenal | [36] |
| 4 | 28.2 | 55.3 | 568.6 | 476.4 | 550.4 | 458.4 | 415.4 | Lutein | [27,37,38] |
| Zeaxanthin | [38,39,40] | ||||||||
| 5 | 32.1 | 37.6 | 621.7 | 381.3 | 515.4 | 399.3 | 529.4 | Unknown | |
| 598.5 | 506.1 | 540.3 | 448.2 | 429.3 | 3,4-dihydrospirilloxanthin * | [41] | |||
| 338.4 | 321.2 | 303.2 | 162.7 | Unknown | |||||
| 6 | 34.2 | 2.5 | 598.6 | 506.0 | 540.3 | 448.2 | 429.3 | 3,4-dihydrospirilloxanthin * | [41] |
| 568.4 | 476.3 | 549.4 | 546.5 | 509.5 | Lutein | [27,37,38] | |||
| Zeaxanthin | [38,39,40] | ||||||||
| 7 | 37.3 | 3.0 | 598.7 | 540.3 | 506.3 | 429.3 | 448.1 | 3,4-dihydrospirilloxanthin * | [41] |
| 567.8 | 550.5 | 509.6 | 529.8 | 461.3 | Hydroxyechinenone | [33] | |||
| 457.3 | 345.1 | 232.7 | 398.3 | 438.2 | Unknown | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pradel, P.; Calisto, N.; Navarro, L.; Barriga, A.; Vera, N.; Aranda, C.; Simpfendorfer, R.; Valdés, N.; Corsini, G.; Tello, M.; et al. Carotenoid Cocktail Produced by An Antarctic Soil Flavobacterium with Biotechnological Potential. Microorganisms 2021, 9, 2419. https://doi.org/10.3390/microorganisms9122419
Pradel P, Calisto N, Navarro L, Barriga A, Vera N, Aranda C, Simpfendorfer R, Valdés N, Corsini G, Tello M, et al. Carotenoid Cocktail Produced by An Antarctic Soil Flavobacterium with Biotechnological Potential. Microorganisms. 2021; 9(12):2419. https://doi.org/10.3390/microorganisms9122419
Chicago/Turabian StylePradel, Paulina, Nancy Calisto, Laura Navarro, Andrés Barriga, Nicolás Vera, Carlos Aranda, Robert Simpfendorfer, Natalia Valdés, Gino Corsini, Mario Tello, and et al. 2021. "Carotenoid Cocktail Produced by An Antarctic Soil Flavobacterium with Biotechnological Potential" Microorganisms 9, no. 12: 2419. https://doi.org/10.3390/microorganisms9122419