
Pteridium aquilinum (Dennstaedtiaceae), a Novel Hyperaccumulator Species of Hexavalent Chromium
by
 , and
, and
Felipe de Jesús Eslava-Silva
1,
María Eugenia Muñíz-Díaz de León
2 and
Manuel Jiménez-Estrada
3,* 1
Posgrado Ciencias Biomédicas, Instituto de Química, Universidad Nacional Autónoma de Mexico, Ciudad de Mexico 04510, Mexico
2
Facultad de Ciencias, Universidad Nacional Autónoma de Mexico, Ciudad de Mexico 04510, Mexico
3
Instituto de Química, Universidad Nacional Autónoma de Mexico, Ciudad de Mexico 04510, Mexico
*
Author to whom correspondence should be addressed.
Appl. Sci. 2023, 13(9), 5621; https://doi.org/10.3390/app13095621
Submission received: 30 March 2023
/
Revised: 21 April 2023
/
Accepted: 28 April 2023
/
Published: 3 May 2023
(This article belongs to the Section Environmental Sciences)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Hexavalent chromium (Cr VI) is a highly toxic environmental pollutant produced as a byproduct of stainless steel manufacture and leather tanning. Several vegetal Cr VI-accumulating species have been investigated in phytoremediation, a promising technology to remove heavy metals from soils and water bodies. The aim of this work was to test the sensitivity, accumulation and remotion of Cr VI in both life cycle phases of the fern Pteridium aquilinum. Both gametophytes and sporophytes were obtained (in vitro) and evaluated (in vitro and using hydroponics) under controlled temperature, photoperiod and humidity conditions. One-month gametophytes were exposed in vitro to K2Cr2O7 (0, 50, 200, 600 and 800 µM). Four-month sporophytes were exposed to K2Cr2O7 (0, 800, 1600 and 6400 µM) under hydroponic conditions. Both phases were harvested at 24, 28, 72 and 168 h post-exposure, and biomass, chlorophyll content (a and b) and the amount of Cr in tissues and culture medium were tested. The results indicate that both phases of the cycle are not sensitive to Cr VI, since chlorosis and reduction in biomass were not observed. The gametophytes accumulated up to 915 mg of Cr×Kg−1 DW, while the sporophytes accumulated up to 11,854 of Cr×kg−1 DW in the underground parts. The sporophytes showed higher Cr uptake in rhizomes and adventitious roots, and despite having a low translocation index toward the leaves, these reached high concentrations as well (2240 mg Cr×Kg−1 DW). Given the uptake capacity in sporophytes, this fern places within the top five species with highest Cr accumulation, and it may be successfully used in phytoremediation methods.
1. Introduction
Chromium (Cr) is found in a series of oxidation states in the environment, with the trivalent (Cr III) and the hexavalent (Cr VI) being the two most common forms [1]. At low concentrations, Cr VI causes severe toxicity, since it has been identified to have mutagenic and carcinogenic properties. Furthermore, it generates free radicals and reactive oxygen species (ROS). Because of its high solubility in water, Cr VI is very active in the environment, making this metal, derived from steel manufacturing and leather tanning, a dangerous pollutant [2,3]. The pollution of soils and water bodies with Cr is a serious problem in ecosystems, since methods for its transformation into a less toxic form remain unknown [4]. In 2015, the extraction of this metal reached over 27,000 million metric tons worldwide [5]. An alternative to manage the pollution of Cr is through phytoremediation [6,7].
Phytoremediation is the use of plants and rhizosphere-related microorganisms to eliminate, transform or capture toxic metals from ecosystems, including Cr VI [6,7]. Plants employed in this technology must have rapid growth, substantial biomass and deep roots, should be easily harvested and be able to tolerate several metals with efficient accumulation within tissues [6]. Studies in the field of phytoremediation are mainly focused on finding new toxic metal hyperaccumulator species to be successfully employed. The choice of species with potential to be applied in bioremediation processes is made in situ by analyzing the individuals that grow in locations highly polluted by heavy metals, which are included in bioassays [8]. The strategies to propose hyperaccumulator species are mainly based on bioassays under controlled conditions, such as in vitro cultures and hydroponic cultures. Both methods allow us to obtain information more readily than field experiments using the whole plant (root, stems and leaves). Furthermore, the experiments on cultures also allow us to modulate temperature, humidity, pH or photoperiod. Thus, it is possible to characterize the optimal growing parameters required by the studied plant [8].
Regarding Cr VI, a plant is considered a hyperaccumulator if it is able to absorb above 0.1% of dry weight (DW) within its tissues [9] or if the plant concentrates above 0.1% of Cr DW into its leaves [10]. Currently, 70 known species of plants can absorb Cr; however, 90% of them fail to accumulate enough metal and cannot be considered hyperaccumulators, so these species do not meet the phytoremediation criteria [4]. Thus, the identification and description of new hyperaccumulator plants is crucial in the remediation of pollution through this biotechnological technique [4].
Ferns are a promising group of plants in phytoremediation. Their life cycle shows alternations of heteromorphic and independent generations [11]: (I) a gametophyte phase with short-term prothallium that produces gametes and (II) a dominant sporophyte phase, represented by a corm constituted by root, stem and leaves with sporangia where meiospores are contained. When these meiospores germinate, they generate the prothallium [12]. In several fern species, it has been observed that both phases are hyperaccumulators and highly tolerant to heavy metals such as lead (Pb), arsenic (As) and Cr from polluted soils and water bodies [13,14,15].
Pteridium aquilinum (L.) Kuhn is a fern seconded to the Dennstaedtiaceae family, and it is a heliophile species and cosmopolite. It successfully grows in highly polluted ecosystems with nickel (Ni) and Cr [16,17]. The presence of this plant in such habitats suggests it has the potential to be used in phytoremediation [18]. However, studies on its ability to accumulate Cr are lacking. Thus, the aim of this work was to determinate the accumulation capacity of P. aquilinum in both gametophytes and sporophytes under controlled conditions by using both in vitro and hydroponic cultures.
In order to characterize the accumulation capacity of Cr VI, both phases in the life cycle of this fern were exposed to increasing concentrations of the metal for different periods of time. The quantification of Cr VI was assessed through atomic absorption spectroscopy (ASS) using the flame method. The results indicate high accumulation from both sporophytes and gametophytes (11,973.93 mg Cr×Kg−1 DW; 915 mg Cr×Kg−1 DW, respectively), where rhizomes and adventitious roots from sporophytes were the organs that showed the highest efficiency of accumulation, approximately 10-fold higher than the gametophytes. This taxon exhibits peculiar ecological and biochemical properties, such as having a considerable vegetative propagation, being an allelopathic, pioneer and heliophile plant and growing in highly perturbed and/or polluted zones. Considering these features along with the data obtained, this species meets the hyperaccumulator criteria and could be considered as a serious biotechnological tool in phytoremediation processes.
2. Materials and Methods
2.1. Biological Material and Propagation
Spores of P. aquilinum were collected at La Cantera (20°39′54.3″ N, 98°39′56.2″ W, 2036 msnm), located in Zacualtipan, Hidalgo, Mexico. The fertile pinnae were dehydrated at 40 °C using an electric drier, then they were sieved and spores were obtained as described by Muñiz et al. [19] and kept at 4 °C until processing.
Young gametophytes and sporophytes were obtained from in vitro cultures. Briefly, 90 mg of spores were washed by shaking in 0.3% NaClO for 2 min. The suspension containing the spores was filtered and washed five consecutive times in sterile water; then, the spores were resuspended in 10 mL of water and 0.25 mL of this suspension was sowed in 10 cm sterile Petri dishes containing ½ MS medium [20]. Fifteen days later, the resultant gametophytes were transplanted to fresh medium, and they were watered twice a week with 5 mL of sterile deionized water until the sporophytes were observed. Between days 60 and 70, the sporophytes were transplanted into perlite pots and watered with a macronutrient solution of ½ MS medium, pH 5.7 [21]. For acclimatization, the humidity was gradually lowered in greenhouse conditions for 30 days.
2.2. Cr Bioassays
To evaluate the accumulation activity of P. aquilinum, the Cr VI absorbed (vs. free in medium) biomass and chlorophyll (a/b) content were determined in both sporophytes and gametophytes treated as follows: 1 g of 30-day-old gametophytes was transplanted into sterile Petri dishes (66 × 15 mm) containing 3 mL of hydroponic sterile solution (½ MS) and then exposed to 0, 50, 200, 600 and 800 µM of K2Cr2O7 (n = 3–5). The sporophyte assays were carried out using 4-month-old plants in perlite pots watered with 100 mL of hydroponic solution and exposed to 0, 800, 1600 and 6400 µM of K2Cr2O7 (n = 3–5). Both phases were exposed to K2Cr2O7 for 24, 48, 72 and 168 h and then harvested for analysis. Three to five independent replicates were made for every concentration and sampling time.
2.3. Chlorophyll Content
The chlorophyll a and b contained in gametophytes and the early leaves were quantified according to the method of Yu et al. [22]. Once the plants were harvested, 0.5 g of fresh tissue was sectioned in small fragments and 80% acetone was added to obtain a final volume of 25 mL. Samples were shaken and protected from light for 24 h, and colorimetric spectrometry was used to evaluate chlorophyll a and b at 645 nm and 663 nm, respectively. The following formula was employed to calculate the chlorophyll content [23]:
where Ca is the chlorophyll a concentration (mg•g−1 FW), Cb is the chlorophyll b concentration (mg•g−1 FW), D is the optical density obtained for the wavelength, V is the final volume (mL), P is the fresh tissue mass (g) and d is the distance the light traveled from the source to the sample (1 cm).
2.4. Quantification Cr VI in Culture Medium and Total Cr Content in Plant Tissue and Media Culture
The Cr VI quantification was carried out using the colorimetric reaction with 1,5-diphenylcarbazide (DFC), since this oxidation state of Cr is spontaneously reduced to Cr III [24]. Aliquots of the culture medium were mixed with DFC (1 volume of 100 mg DFC in 95% ethanol and 1 volume of 10% H2SO4) and the absorbance of the mixture was measured at 540 nm (UV-5100B spectrophotometer, METASH, Shanghai, China). DFC specifically reacts with Cr VI species but not with Cr III; interference from some structural chemical analogues, such as molibdate, telurate and vanadate, or from iron may occur. Then, to avoid an overestimation of Cr VI, control reactions of DFC with fresh culture medium were routinely conducted. For calibration, a commercial stock 0.1% Cr VI solution (Sigma, México city, Mexico) was used as standard. The reaction was linear up to 50 nmol Cr VI (24).
For the total Cr quantification, once the tissue samples were exposed to different concentrations of K2Cr2O7, both young gametophytes and sporophytes were washed with ethylenglycol-bis (2-aminoether)-N,N,N′,N′-tetraacetic acid (EGTA) 1 mM pH 7.0, then with 20% ethylendiaminotetraacetic acid (EDTA) pH 7.0. The remaining Cr absorbed in the plant surface was eliminated using a solution of 5% nitric oxide (NO) and distilled water afterwards. Then, the samples were dried at 60 °C and subsequently homogenized with liquid nitrogen and kept at room temperature until determination.
The Cr quantification in tissues as well in the substrates of gametophytes and sporophytes (water and perlite, respectively) was assessed following the United States of America EPA 3052, 3051 and 3015 methods [25,26,27,28]. The Cr was quantified through ASS (SpectrAAA 880, Varian, CA, USA) according to the method of Santillán et al. [29]. The instrument was calibrated using a Cr standard curve (1, 2, 3, 4 and 5 mg•Kg −1) (TraceCERT®, 1000 mg Cr•l−1 in NO, N° Cat. 02733-250ML). For validation of the Cr extraction, a standard vegetal material was employed (Standard Reference Material® 1573a, National Institute of Standards and Technology) by obtaining 90 to 95% of Cr extraction.
Once the Cr values were obtained from the samples, the following parameters were determined:
The translocation percentage for sporophytes was calculated through the following formula [30]:
For both sporophytes and gametophytes, the removal percentage was calculated using the following formula [31]:
where C is the Cr concentration (mg•Kg−1) obtained from the plant structures and substrates.
2.5. Statistical Analysis
For statistical tests, two-way analysis of variance (two-way ANOVA) was employed, followed by a Bonferroni test. Values of p < 0.05 were considered to be statistically significant different. Data were analyzed using the Prims 6 software (Graphpad, San Diego, CA, USA, EE.UU).
3. Results
3.1. Quantification of Cr VI in Hydroponic Culture Medium
Since Cr VI is spontaneously reduced to Cr III in the presence of organic matter, first, the reduction rate was determined under in vitro conditions using the culture medium containing the gametophytes or sporophytes, as well as the cultures alone. The results showed that Cr VI is found in the medium throughout the entire assay (Tables S1 and S2).
3.2. Sensitivity of P. aquilinum to Cr
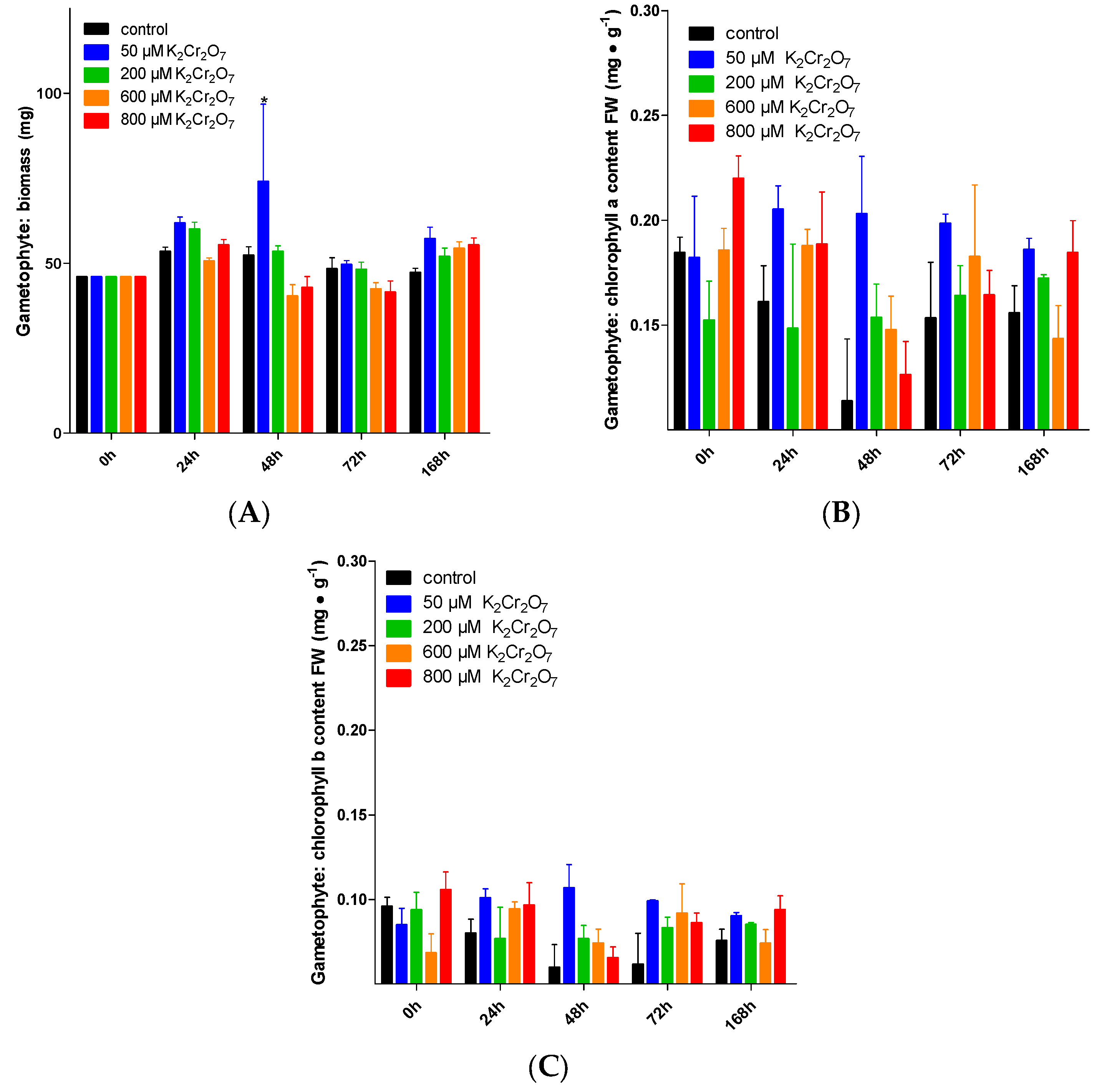
The vegetal species when exposed to Cr VI exhibited evident signs of toxicity, such as lower biomass and chlorosis (lower quantities of chlorophyll a and b) [1,8]. Here, such parameters were tested in treated gametophytes and sporophytes. Gametophytes were not sensitive to the Cr VI treatments (from 0, 50, 200 and 600 µM K2Cr2O7), since biomass remained constant (approximately 52 mg DW). Only a biomasas with K2Cr2O7 50, 200 µM at 48 h was an increment observed. However, such differences were not observed in subsequent times when compared to the control, and they persisted until the end of the treatment (Figure 1A). On the other hand, a and b remained at ~0.167 and ~0.084 mg•g−1 FW, respectively (Figure 1B,C).
Increases in the biomass of leaves from sporophytes were observed after 168 h when treated with K2Cr2O7 800 and 6,400 µM, with a significant increase compared to the control: 447.82 mg DW and 427.32 DW, respectively (Figure 2A). On the other hand, for the biomass of underground parts (rhizomes and adventitious roots) at the end of the experiment, an increase was observed in the metal-exposed plants, with K2Cr2O7 800 µM rising to 165.06 mg of biomass, while with K2Cr2O7 6,400 µM, the biomass was 155.7 mg (Figure 2B). Regarding chlorophyll a and b content in leaves, there were no significant changes with either K2Cr2O7 800 µM or 1600 µM. Regarding the chlorophyl a content, despite the increase observed at 48 h when exposed to 6400 µM K2Cr2O7 (2.118 mg•g−1 FW), at the end of the experiment, no significant differences were observed when compared to the control (Figure 2C,D).
3.3. Cr Uptake
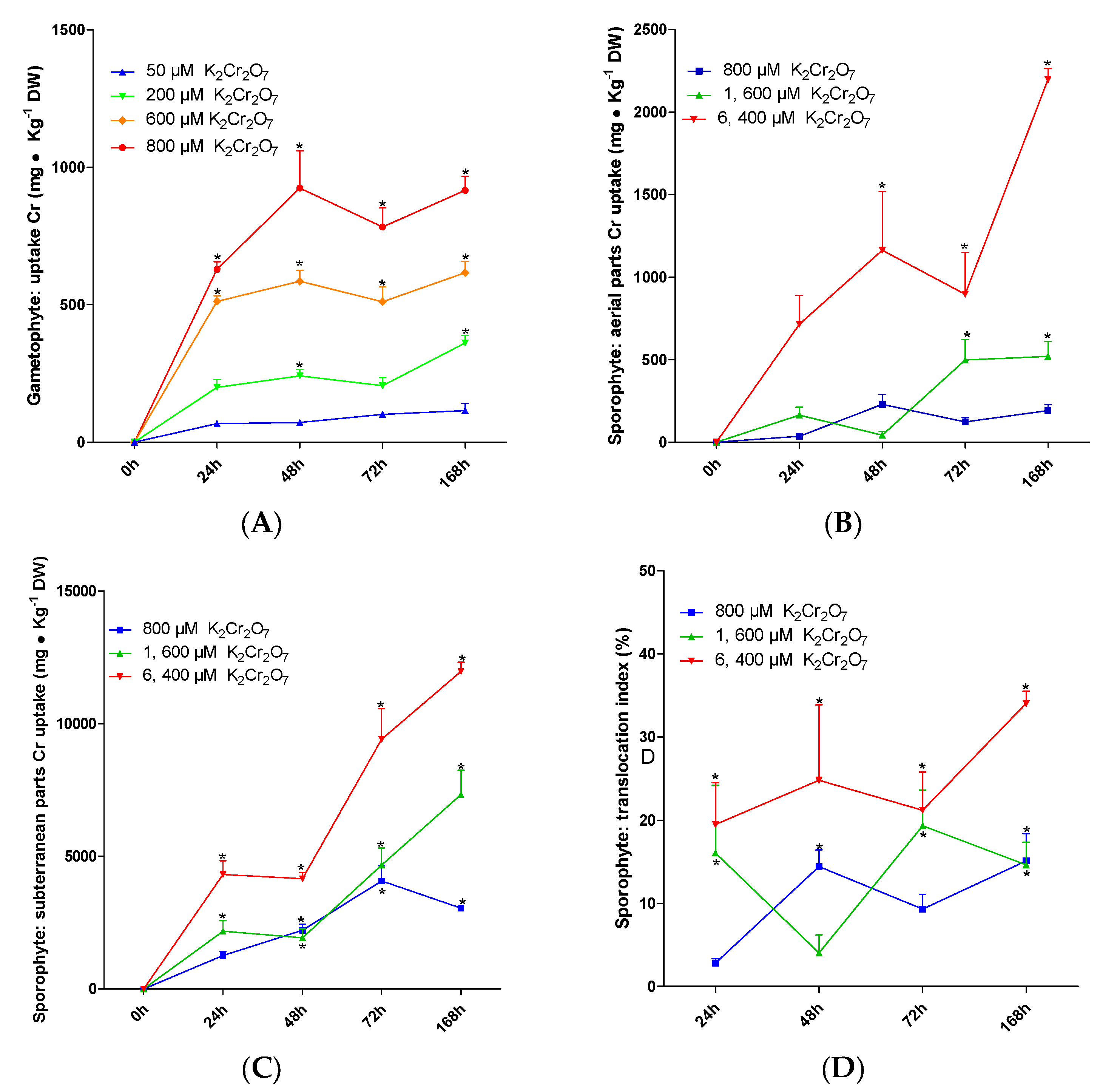
To consider the use of a species in phytoremediation, one of the main criteria is its capacity to accumulate high amounts of the metal. The results from these experiments showed that P. aquilinum can accumulate up to 915 mg Cr •kg−1 DW when treated with K2Cr2O7 800 µM for 168 h. On the other hand, the K2Cr2O7 treatments from 50 to 600 µM produced a maximum absorption at 24 h. The absorptions obtained with the different concentrations are as follows: 116 mg Cr×Kg−1 DW with K2Cr2O7 50 μM, 362 mg Cr×Kg−1 DW with K2Cr2O7 200 μM and 616 mg Cr×Kg−1 DW with K2Cr2O7 600 μM (Figure 3A).
The maximum Cr accumulated by the sporophytes was induced by K2Cr2O7 6400 µM at 168 h. The aerial parts absorbed 2198.132 mg Cr×Kg−1 DW (Figure 3B), while the underground parts absorbed 11,973.93 mg Cr×Kg−1 DW (Figure 3C). For every treatment, the underground parts showed the highest capacity; with K2Cr2O7 800 and 1600 µM, the absorption was 3,051.899 and 7,734 mg Cr×Kg−1 DW, respectively (Figure 3C).
The sporophytes showed a low percentage of translocation from the underground parts to the aerial parts. The highest value was observed in plants exposed to K2Cr2O7 6400 µM at 168 h (34.048% of total Cr accumulated). This effect was observed in the aerial parts. The individuals exposed to K2Cr2O7 800 µM achieved their highest translocation percentage (15.098%) at 72 h, and with the 1600 µM treatment at 168 h, the percentage was 14.608% (Figure 3D).
The remotion percentage represents the amount of Cr eliminated from the substrate by the plant. The gametophytes showed a maximum remotion percentage of 47.8% when treated with K2Cr2O7 50 µM for 168 h. The remotion percentage was lower than 30% for the remaining treatments with K2Cr2O7 (Figure 4A). On the other hand, the highest remotion percentages in the sporophytes were 90.414% and 90.802% at 168 h with K2Cr2O7 800 µM and 1600 µM, respectively (Figure 4B). In addition, when exposed to K2Cr2O7 6400 µM, the Cr remotion was slightly lower, showing 83.983% efficiency Figure 4B.
4. Discussion
4.1. Application of Culture In Vitro and Hydroponics to Find New Hyperaccumulator Species
Bioassay applications with control of most of the variables, such as in vitro and hydroponic cultures, have served to evaluate the tolerance, accumulation and translocation of metals in plants under optimal conditions. Thus, it is possible to evaluate the maximum resistance and accumulation of the organs from plants growing in metal-polluted sites. The bioassays on these taxa have generated relevant information about their use in the phytoremediation field. Examples of these strategies are the works using species from the genera Salix L., Populus L. and Picea A. Dietr. [8]. Because of the small size of gametophytes from P. aquilinum, the experiments in these structures were carried out on in vitro cultures. Meanwhile, as the sporophytes have a considerably larger size, they were kept in hydroponic culture. This enabled the determination of the maximum resistance and accumulation of Cr VI, as well as the opportunity to establish whether it fulfils the required criteria to be used on polluted sites and is able to remove the metal.
4.2. Sensitivity of P. aquilinum to Cr
According to the data, both gametophytes and sporophytes from P. aquilinum showed high resistance to several concentrations of Cr VI, since neither chlorosis nor biomass reduction were observed after the exposure to this metal. This effect was enhanced in the underground parts of sporophytes since biomass increased. Although it has been documented that Cr is a micronutrient to plants that stimulates both parameters, its concentrations should not exceed 1 µM, since this metal may induce toxicity [2,32]. In this work, the P. aquilinum plants were exposed to higher concentrations of Cr and such structures showed no signs of damage from toxicity.
Comparing the P. aquilinum sensitivity, specifically the of gametophytes, with other species of seed plants showing resistance to Cr VI is difficult since the tissue structure is simple (thalloid) when contrasted with seed plants, which possess a vascular system (corm). However, the sensitivity shown by this phase may be compared to assays on germinating seeds exposed to Cr VI, since this is the most vulnerable phase in the life cycle of spermatophytes. In order to compare our results from gametophytes to other works, molar concentrations were converted to parts per million (ppm), where K2Cr2O7 VI 50 µM equals 4.1 ppm. Thus, K2Cr2O7 VI 200 µM = 24.3 ppm, 600 µM = 30.9 ppm and 800 µM = 37.1 ppm. Although the P. aquilinum gametophytes lack structural complexity, they did not show any sign of damage or structural deterioration after being exposed to different concentrations of K2Cr2O7. The opposite has been shown in several studies, where treatments with even lower concentrations of Cr VI during germination negatively impacted the chlorophyll a and b and biomass of Salvia sclarea L. (1, 5 and 10 ppm) [33], Echinochloa colona (L.) Link. (1.25 and 2.5 ppm) [34], Caesalpinia pulcherrima (L.) Sw. (25 ppm) [35], Zea mays L. (5 and 10 ppm) [36], Lycopersicon esculentum Mill. (5 and 10 ppm) [36] and Brassica oleracea L. (5 and 10 ppm) [36]. Because of their tissular simplicity, it is expected to observe a higher sensitivity of gametophytes to Cr VI than that of a plant with more complex tissues and organs (such as roots, stems and leaves), even in young stages (e.g., germination), in contrast to our results. Thus, the homeostasis systems in gametophytes are likely to be more efficient. However, this remains to be studied in future works.
Regarding the results from roots, rhizomes and leaves from sporophytes exposed to Cr VI, the data indicate that leaves did not show chlorosis or biomass reduction, whereas rhizomes and adventitious roots increased their size. The effect on both parameters suggests that the amount of Cr VI used in this work not only did not exceed the plant’s capacity to resist the toxicity, but it also promoted the growth of underground parts. In fact, this has not been reported previously in other species exposed to Cr VI. Thus, P. aquilinum is one of the most resistant species to Cr reported in the literature to date. This fern is even more resistant than Convolvulus arvensis L. [37], Helianthus annuus L. [38], Prosopis sp. L. [39] and Datura innoxia Mill. [40], all of which are reported to be highly resistant plants to Cr with superior accumulation capacity. However, it has been reported that such species show a few toxicity signs when challenged with higher concentrations of the metal.
4.3. P. aquilinum Cr Uptake
The sporophytes from P. aquilinum accumulate high amounts of Cr VI in their tissues with respect to other terrestrial ferns such as Pellaea falcata R.Br. (35 mg Cr×Kg−1 DW in fronds and 130 mg Cr×Kg−1 DW in roots) [41] and Pteris vittata L. (5717 mg Cr×Kg−1 DW in rhizome-roots) [42]. In fact, its capacity is similar to the aquatic ferns [43] Azolla filiculoides Lam. (12,383.6 mg Cr×Kg−1 DW), A. microphylla Kaulf. (14,931.7) and A. pinnata (R.) Brown (9125.3 mg Cr×Kg−1 DW), all of which are employed in water body phytoremediation. Regarding the terrestrial species, this fern surpasses flowering plants such as Datura innoxia (10 469 mg Cr×Kg−1 DW in roots) [40], Helianthus annuus (11,030 mg CrKg-1 DW in roots) [38] and Prosopis sp. (10,983 mg Cr×Kg−1 DW in roots) [39]. The accumulation capacity of P. aquilinum is only surpassed by Convolvulus arvensis, which accumulates an average of >20,000 mg Cr×Kg−1 DW [37]. The biological relevance of Cr VI accumulation in plants remains unknown [5], however, the fern P. vittata accumulates As, it is proposed that the plant uses this metal as strategy against the herbivory [42], this remains to be elucidated for the Cr VI.
The sporophytes from P. aquilinum accumulate large quantities of Cr VI in their tissues, surpassing plants such as Datura innoxia (10,469 mg Cr×Kg−1 DW in roots) [40], Helianthus annuus (11,030 mg Cr×Kg−1 in roots) [38] and Prosopis sp. (10,983 mg Cr×Kg−1 DW in roots) [39]. Likewise, when compared to Pteris vittata L. (5717 mg Cr×Kg−1 DW in rhizomes and roots) [42], the only terrestrial fern in which the accumulation capacity has been tested, P. aquilinum is more efficient. It is worth highlighting that according to the data, P. aquilinum is surpassed only by Convolvulus arvensis, which accumulates on average >20,000 mg Cr×Kg−1 DW in its roots [37].
Bioassays describing the absorption of different oxidation states of Cr in fern gametophytes are lacking; thus, it is not possible to compare the accumulation capacity of gametophytes from P. aquilinum to other plants from these taxa in this phase of the life cycle. Only a few studies report the accumulating capacity of toxic metals in this group of plants, i.e., Athyrium yokoscense (Franch. & Sav.) Christ absorbs ~5500 mg Pb•Kg−1 DW, ~7500 mg Cd•Kg−1 DW [44] and 443 mg Ba•Kg−1 DW [45], Pityrogramma calomelanos (L.) Link accumulates 646.51 mg Pb•Kg−1 DW and 1885 mg Pb•Kg−1 DW [46] and Pteris vittata 2180 mg As•Kg−1 DW [47]. It is worth highlighting that gametophytes with metal accumulation retain a correspondence with the sporophytes, since a higher accumulation for sporophytes versus gametophytes has been reported. Such an effect was also observed in P. aquilinum, where the sporophyte accumulation was approximately 13-fold higher than that of the gametophytes, suggesting the presence of similar mechanisms in both life cycle phases despite the differences between them. Thus, the gametophyte may be proposed as a metal-indicating plant, whether tolerance and accumulation bioassays in this phase were developed and show resistance and accumulation, it is probable that the sporophyte show high performance for the evaluated metals.
The accumulating curve from the assays on gametophytes showed that metal absorption increases with time, reaching the asymptote of the curve, the time that determines the maximum capacity of Cr VI accumulation, at 48 h. For the sporophytes, the highest accumulation was not determined (as observed in Figure 3B,C) since in both curves from leaves, rhizomes and adventitious roots, the asymptote is not observed, suggesting that higher accumulation may be achieved. It is possible that by exposing the sporophytes for longer periods of time and increasing the concentration of Cr, a higher accumulation may be observed. The analysis of Cr VI absorption using young sporophytes (~4 months) suggests that the mature plants (with reproductive structures) could accumulate higher amounts of Cr VI.
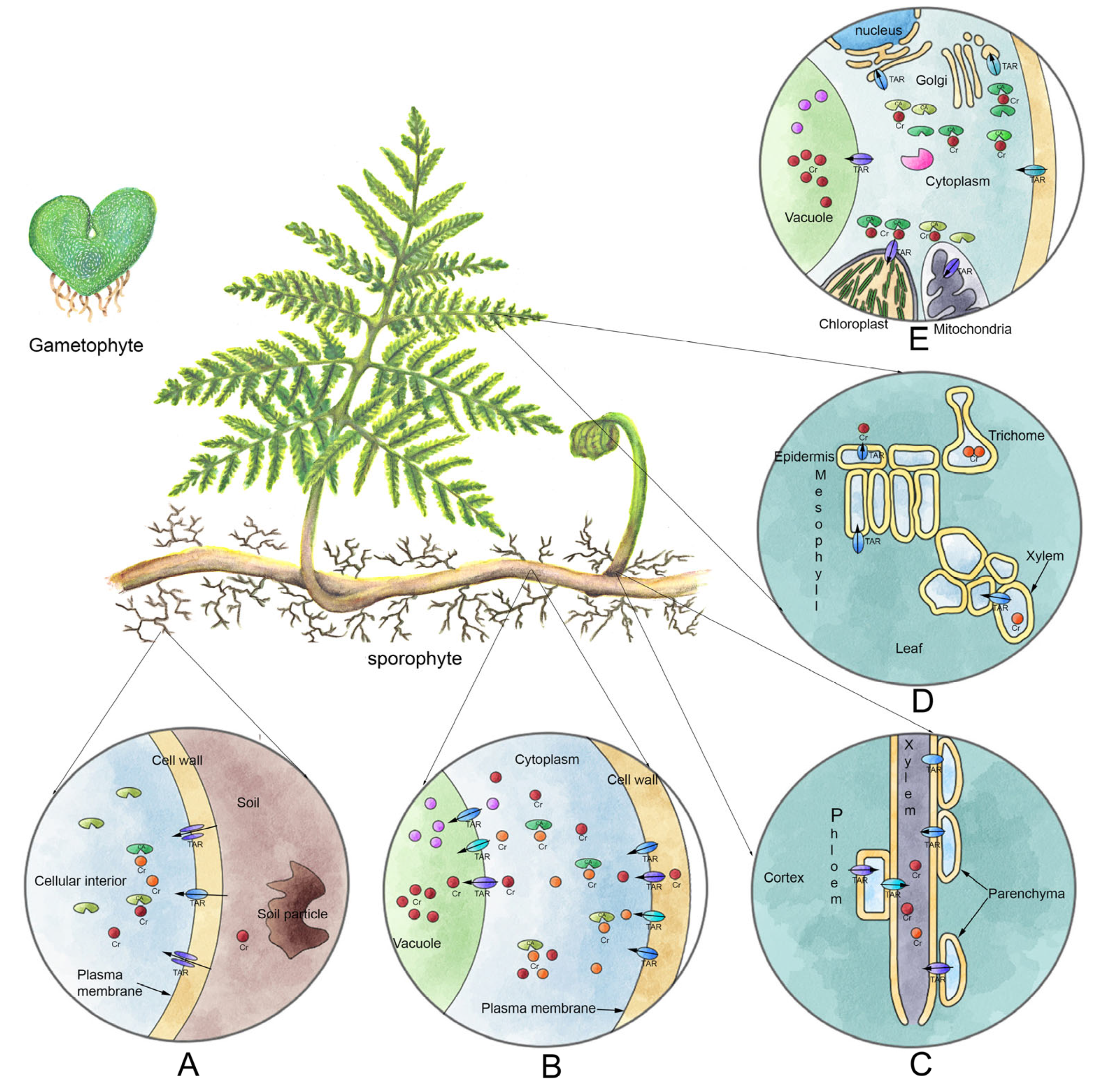
The low translocation percentage shown by P. aquilinum (<50%) could be due, as reported by Hayat et al. [5], to the Cr VI entry via symplast to absorption organs (rhizomes/roots). By making contact with organic matter, it reduces to Cr III. This form has low solubility and is mobilized into vacuoles, and only a small fraction is transported to aerial parts. This modality of distribution has been extensively reported in plants exposed to Cr salts [22,30,37,38,39,40,42,48]. Such a pathway may explain the lower accumulation values observed in leaves, since the highest amount of metal was accumulated in rhizomes/roots (Figure 5). However, it is worth highlighting that the leaves accumulated 2198 mg Cr×Kg−1 DW, a two-fold higher efficiency than that reported for other species [49,50] with values under 1000 mg Cr×Kg−1 DW.
4.4. P. aquilinum as Novel Cr Hyperaccumulator Species
Sporophytes from P. aquilinum showed the highest Cr VI absorption values. It also fulfills most of the criteria to be considered a hyperaccumulator. Both roots and rhizomes accumulated an average of 1.1973% Cr DW, and 0.2198% Cr DW was accumulated in leaves, significantly higher than the values previously reported by Morel et al. [51], Branquinho et al. [52] and Reeves and Baker [10]. Such works report that to be considered a hyperaccumulator, a given species must absorb a minimum percentage of 0.1% DW of Cr. The roots and rhizomes from P. aquilinum accumulated 11,973.93 mg Cr×Kg−1 DW, while the reported values for Cr-absorbing plants (under natural conditions) range from 0.2 to 5 mg Cr×Kg−1 DW. Our results indicate that P. aquilinum has an efficiency about 50,000-fold higher. Thus, this fern certainly could be considered a hyperaccumulator.
Moreover, the criteria reported by Baker and Walker [49] and Baker et al. [50] establish that a hyperaccumulator plant must incorporate higher amounts of Cr in the aerial parts (stem and leaves) than the roots. With such criteria, the fern studied here could not be considered a hyperaccumulator. It is worth noting that a consensus rule to classify hyperaccumulator plants is lacking. Here, three different criteria are cited and P. aquilinum fulfills two of them. The third criterion proposed by Baker and Walker [49] and Baker et al. [50] could be discarded since there are no reports of any plant capable of absorbing higher amounts of Cr in leaves than roots, possibly because of its low translocation rate.
The capacity of gametophytes to remove Cr VI from culture media, regardless of its tissular simplicity, surpasses that exhibited by vascular plants, such as Nymphaea spontanea K.C. Landon, (31.6%) [53], Spirodela punctata (G.F.W. Meyer) Thomps. (19%) [54], Spirodeia polyrhiza (L.) Schleiden 15% [54] and Lemna gibba L. 10% [54]. On the other hand, the sporophytes by far surpassed the remotion capacity shown by the gametophytes. This could be compared to Eichhornia crassipes (Mart.) Solms [55], which removes 84% of metal, Spirodela polyrhiza (L.) Schleid 75% [56], Salvinia molesta D.S. Mitch. 90% [56] and Miscanthus Sinensis 90–92% [57]. These plants have been reported to have the highest remotion values. It is worth highlighting that sporophytes from P. aquilinum have terrestrial habits and showed similar or even higher remotion values than those shown by aquatic angiosperms.
The sporophytes from P. aquilinum exhibited Cr VI hyperaccumulation and robust resistance. Moreover, this species has interesting biological attributes; it is cosmopolite [58,59,60] and has high underground propagation, fire resistance [61] and drought resistance [62]. Altogether, these characteristics allow it to establish and grow in perturbed and polluted soils. Furthermore, interesting metabolic traits have been reported in this species; it has allelopathic effects [63] and several defenses against herbivory [64], both important parameters considered in large-scale ex situ remediation processes. The allelopathy prevents other species from colonizing the bioreactor where the remediation process is carried out. Meanwhile, the anti-predatory strategies prevent biomagnification [6], namely the entry of Cr into the trophic network when it has been accumulated in the vegetal tissues. In consequence, and taken together with previously reported biological features, P. aquilinum is an excellent candidate due to its capacity for accumulation, remotion and resistance to Cr VI characterized for the first time in this work.
5. Conclusions
This research showed the accumulation capacity and tolerance to Cr VI in organs from the two phases of the P. aquilinum life cycle. The sporophyte is the longest phase and has the largest size as well, thus, it can be identified in several vegetations. Concluding and according to the results presented here, it can be considered as a new hyperaccumulator species for Cr VI. Its high absorption capacity places this fern among the five top accumulator species studied so far. Because of its accumulation capacity for, removal of and tolerance to Cr VI, as well as its low translocation rate, pioneering nature, anti-predatory strategies, resistance to perturbation, fire resistance, large biomass and high vegetative propagation, this plant is an excellent candidate for phytoremediation. Once the bioremediatory potential of P. quilinum is characterized, it is necessary to study containment methods to prevent invasion toward surrounding ecosystems that interact with the polluted area subjected to remediation.
As mentioned above, the highest values of tolerance and absorption of Cr VI were observed in the sporophytes. However, the gametophytes showed relatively high values as well. These data, taken together, represent an experimental advantage, since ferns had never been considered for remediation before. The gametophyte, being the shorter phase with lower structural complexity (no deep-root system) and low biomass (barely a few mm in size), does not fulfill the requirements to be used in phytoremediation. Nonetheless, its sensitivity and Cr absorption suggest its potential use as a bioindicator of the metal. It would be convenient to study its tolerance to different metals, thus using it for pollutant monitoring.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app13095621/s1, Table S1: Quantification of Cr VI in hydroponic culture medium in gametophytes; Table S2: Quantification of Cr VI in hydroponic culture medium in sporophytes.
Author Contributions
F.d.J.E.-S. designed and conducted all the experiments of this work and also wrote the manuscript draft. M.E.M.-D.d.L. helped with the fern culture and propagation and wrote the manuscript draft. M.J.-E. provided the reagents and place to carry out this investigation and wrote the manuscript draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Felipe de Jesús Eslava-Silva is grateful to the PhD program in Ciencias Biomédicas of Universidad Nacional Autónoma de México (UNAM) and CONACyT for providing scholarship number 47165. Rafael Moreno Sánchez and Jorge Donato García García from Instituto Nacional de Cardiología, Departamento de Bioquímica, for their important comments during the development of this project.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper. All authors approved the final manuscript and agreed to submission.
References
- Cervantes, C.; Campos-García, J.; Devars, S.; Gutiérrez-Corona, F.; Loza-Tavera, H.; Torres-Guzmán, J.C.; Moreno-Sánchez, R. Interactions of chromium with microorganisms and plants. FEMS Microbiol. Rev. 2001, 25, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Khalique, G.; Irfan, M.; Wani, A.S.; Tripathi, B.N.; Ahmad, A. Physiological changes induced by chromium stress in plants: An overview. Protoplasma 2012, 249, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Nirola, R.; Megharaj, M.; Subramanian, A.; Thavamani, P.; Ramadass, K.; Aryal, R.; Saint, C. Analysis of chromium status in the revegetated flora of a tannery waste site and microcosm studies using earthworm E. fetida. Environ. Sci. Pollut. Res. 2018, 25, 5063–5070. [Google Scholar] [CrossRef] [PubMed]
- Sarma, H. Metal hyperaccumulation in plants: A review focusing on phytoremediation technology. J. Environ. Sci. Technol. 2011, 4, 118–138. [Google Scholar] [CrossRef]
- Shahid, M.; Shamshad, S.; Rafiq, M.; Khalid, S.; Bibi, I.; Niazi, N.K.; Dumat, C.; Rashid, M.I. Chromium speciation, bioavailability, uptake, toxicity and detoxification in soil-plant system: A review. Chemosphere 2017, 178, 513–533. [Google Scholar] [CrossRef]
- Arthur, E.L.; Rice, P.J.; Rice, P.J.; Anderson, T.A.; Baladi, S.M.; Henderson, K.L.; Coats, J.R. Phytoremediation—An overview. Crit. Rev. Plant. Sci. 2005, 24, 109–122. [Google Scholar] [CrossRef]
- Antoniadis, V.; Levizou, E.; Shaheen, S.M.; Ok, Y.S.; Sebastian, A.; Baum, C.; Prasad, M.N.V.; Wenzel, W.W.; Rinklebe, J. Trace elements in the soil-plant interface: Phytoavailability, translocation, and phytoremediation—A review. Earth Sci. Rev. 2017, 171, 621–645. [Google Scholar] [CrossRef]
- Capuana, M. Heavy metals and woody plants-biotechnologies for phytoremediation. iForest 2011, 4, 7–15. [Google Scholar] [CrossRef]
- Buscaroli, A. An overview of indexes to evaluate terrestrial plants for phytoremediation purposes. Ecol. Indic. 2017, 82, 367–380. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J. Metal-accumulating plants. In Phytoremediation of Toxic Metals: Using Plants to Clean Up the Environment, 1st ed.; Raskin, I., Ensley, B.D., Eds.; John Wiley and Sons: New York, NY, USA, 2000; pp. 193–229. [Google Scholar]
- Muñiz, M.E.; Pérez, J.C.; Eslava, F.; Jiménez, K.; Valencia, A. Atlas Pteridológico de la Zona Núcleo Poniente de la Reserva Ecológica del Pedregal de San Ángel, 1st ed.; Las Prensas de Ciencias: Ciudad de México, México, 2019; pp. 13–19. [Google Scholar]
- Eslava-Silva, F.D.J.; Jiménez-Durán, K.; Jiménez-Estrada, M.; Muñiz-Díaz-de-León, M.E. Morfo-anatomía del ciclo de vida del helecho Pteridium aquilinum (Dennstaedtiaceae) en cultivo in vitro. Rev. De Biol. Tropical. 2020, 68, 12–22. [Google Scholar] [CrossRef]
- Bennicelli, R.; Stępniewska, Z.; Banach, A.; Szajnocha, K.; Ostrowski, J. The ability of Azolla caroliniana to remove heavy metals (Hg (II), Cr (III), Cr (VI)) from municipal waste water. Chemosphere 2004, 55, 141–146. [Google Scholar] [CrossRef]
- Gumaelius, L.; Lahner, B.; Salt, D.E.; Banks, J.A. Arsenic hyperaccumulation in gametophytes of Pteris vittata: A new model system for analysis of arsenic hyperaccumulation. Plant Physiol. 2004, 136, 3198–3208. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, P.; Singh, J.; Kumar, P. Potential of water fern (Azolla pinnata R. Br.) in phytoremediation of integrated industrial effluent of SIIDCUL, Haridwar, India: Removal of physicochemical and heavy metal pollutants. Int. J. Phytoremediation 2020, 22, 392–403. [Google Scholar] [CrossRef]
- Kamachi, H.; Komori, I.; Tamura, H.; Sawa, Y.; Karahara, I.; Honma, Y.; Naoya, W.; Tokimasa, K.; Kenji, M.; Susumu, I.; et al. Lead tolerance and accumulation in the gametophytes of the fern Athyrium yokoscense. J. Plant Res. 2005, 118, 137–145. [Google Scholar] [CrossRef]
- Praveen, A.; Pandey, V.C. Pteridophytes in phytoremediation. Environ. Geochem. Health. 2020, 42, 2399–2411. [Google Scholar] [CrossRef]
- Kubicka, K.; Samecka-Cymerman, A.; Kolon, K.; Kosiba, P.; Kempers, A.J. Chromium and nickel in Pteridium aquilinum from environments with various levels of these metals. Environ. Sci. Pollut. Res. 2015, 22, 527–534. [Google Scholar] [CrossRef]
- Muñiz-Díaz de León, M.E.; Pérez-García, B.; Márquez-Guzmán, J.; Mendoza-Ruiz, A. Developmental gametophyte morphology of seven species of Thelypteris subg. cyclosorus (Thelypteridaceae). Micron 2008, 39, 351–1362. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Taras, M.J. Standard Methods for the Examination of Water and Wastewater, 13th ed.; American Public Health Association: Washington, DC, USA, 1971; pp. 1–114. [Google Scholar]
- Yu, X.Z.; Gu, J.D.; Xing, L.Q. Differences in uptake and translocation of hexavalent and trivalent chromium by two species of willows. Ecotoxicology 2008, 17, 747–755. [Google Scholar] [CrossRef]
- Maclachlan, S.; Zalik, S. Plastid structure, chlorophyll concentration, and free amino acid composition of a chlorophyll mutant of barley. Can. J. Bot. 1963, 41, 1053–1062. [Google Scholar] [CrossRef]
- García-García, J.D.; Rodríguez-Zavala, J.S.; Jasso-Chávez, R.; Mendoza-Cozatl, D.; Moreno-Sánchez, R. Chromium uptake, retention and reduction in photosynthetic Euglena gracilis. Arch. Microbiol. 2009, 191, 431–440. [Google Scholar] [CrossRef] [PubMed]
- U.S. EPA. Method 3051A: Microwave Assisted Acid Digestion of Sediments, Sludge, Soils, and Oils. Available online: https://www.epa.gov/esam/us-epa-method-3051a-microwave-assisted-acid-digestion-sediments-sludges-and-oils (accessed on 29 March 2023).
- U.S. EPA. Method 3052: Microwave Assisted Acid Digestion of Siliceous and Organically based Matrices. Available online: https://www.epa.gov/hw-sw846/sw-846-test-method-3052-microwave-assisted-acid-digestion-siliceous-and-organically-based (accessed on 29 March 2023).
- U.S. EPA. Method 3015: Microwave Assisted Acid Digestion of Aqueous Simple and Extracts. Available online: https://www.epa.gov/esam/epa-method-3015a-microwave-assisted-acid-digestion-aqueous-samples-and-extracts (accessed on 29 March 2023).
- U.S. EPA. Introduction to Phytoremediation. Available online: EPA/600/R-99/107 (accessed on 29 March 2023).
- Juárez-Santillán, L.F.; Lucho-Constantino, C.A.; Vázquez-Rodríguez, G.A.; Cerón-Ubilla, N.M.; Beltrán-Hernández, R.I. Manganese accumulation in plants of the mining zone of Hidalgo, Mexico. Bioresour. Technol. 2010, 101, 5836–5841. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, M.; Pattnaik, M.M.; Mishra, A.K.; Patra, H.K. Bio-concentration of chromium—An in situ phytoremediation study at South Kaliapani chromite mining area of Orissa, India. Environ. Monit. Assess. 2012, 184, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Saeed, T.; Alam, M.K.; Miah, M.J.; Majed, N. Removal of heavy metals in subsurface flow constructed wetlands: Application of effluent recirculation. Environ. Sustain. Indic. 2021, 12, 100–146. [Google Scholar] [CrossRef]
- Vakili, B.; Karimi, F.; Sharifi, M.; Behmanesh, M. Chromium-induced tropane alkaloid production and H6H gene expression in Atropa belladonna L. (Solanaceae) in vitro-propagated plantlets. Plant Physiol. Biochem. 2012, 52, 98–103. [Google Scholar] [CrossRef]
- Corradi, M.G.; Bianchi, A.; Albasini, A. Chromium toxicity in Salvia sclarea—I. Effects of hexavalent chromium on seed germination and seedling development. Environ. Exp. Bot. 1993, 33, 405–413. [Google Scholar] [CrossRef]
- Rout, G.R.; Samantaray, S.; Das, P. Effects of chromium and nickel on germination and growth in tolerant and non-tolerant populations of Echinochloa colona (L.) Link. Chemosphere 2000, 40, 855–859. [Google Scholar] [CrossRef]
- Iqbal, M.Z.; Saeeda, S.; Shafiq, M. Effects of chromium on an important arid tree (Caesalpinia pulcherrima) of Karachi city, Pakistan. Int. J. Ecol. Probl. Biosph. 2001, 20, 404–413. [Google Scholar]
- Di Toppi, L.S.; Fossati, F.; Musetti, R.; Mikerezi, I.; Favali, M.A. Effects of hexavalent chromium on maize, tomato, and cauliflower plants. J. Plant Nutr. 2002, 25, 701–717. [Google Scholar] [CrossRef]
- Gardea-Torresdey, J.L.; Peralta-Videa, J.R.; Montes, M.; De la Rosa, G.; Corral-Diaz, B. Bioaccumulation of cadmium, chromium and copper by Convolvulus arvensis L.: Impact on plant growth and uptake of nutritional elements. Bioresour. Technol. 2004, 92, 229–235. [Google Scholar] [CrossRef]
- January, M.C.; Cutright, T.J.; Keulen, H.V.; Wei, R. Hydroponic phytoremediation of Cd, Cr, Ni, As, and Fe: Can Helianthus annuus hyperaccumulate multiple heavy metals? Chemosphere 2008, 70, 531–537. [Google Scholar] [CrossRef]
- Aldrich, M.V.; Gardea-Torresdey, J.L.; Peralta-Videa, J.R.; Parsons, J.G. Uptake and reduction of Cr (VI) to Cr (III) by mesquite (Prosopis spp.): Chromate—Plant interaction in hydroponics and solid media studied using XAS. Environ. Sci. Technol. 2003, 37, 859–1864. [Google Scholar] [CrossRef]
- Vernay, P.; Gauthier-Moussard, C.; Jean, L.; Bordas, F.; Faure, O.; Ledoigt, G.; Hitmi, A. Effect of chromium species on phytochemical and physiological parameters in Datura innoxia. Chemosphere 2008, 72, 763–771. [Google Scholar] [CrossRef]
- Kachenko, A.G.; Singh, B.; Bhatia, N.P. Heavy metal tolerance in common fern species. Aust. J. Bot. 2007, 55, 63–73. [Google Scholar] [CrossRef]
- Kalve, S.; Sarangi, B.K.; Pandey, R.A.; Chakrabarti, T. Arsenic and chromium hyperaccumulation by an ecotype of Pteris vittata–prospective for phytoextraction from contaminated water and soil. Curr. Science. 2011, 100, 888–894. [Google Scholar]
- Arora, A.; Saxena, S.; Sharma, D.K. Tolerance and phytoaccumulation of chromium by three Azolla species. World J. Microbiol. Biotechnol. 2006, 22, 97–100. [Google Scholar] [CrossRef]
- Kamachi, H.; Morishita, K.; Hatta, M.; Okamoto, A.; Fujii, K.; Imai, N.; Hiyama, S. Lead and cadmium tolerance and accumulation of proanthocyanidin-deficient mutants of the fern Athyrium yokoscense. Int. J. Plant Biol. 2021, 12, 9330. [Google Scholar] [CrossRef]
- Kamachi, H.; Kitamura, N.; Sakatoku, A.; Tanaka, D.; Nakamura, S. Barium accumulation in the metalliferous fern Athyrium yokoscense. Theor. Exp. Plant Physiol. 2015, 27, 99–107. [Google Scholar] [CrossRef]
- Pulukkunadu Thekkeveedu, R.; Hegde, S. In vitro spore germination and phytoremediation of Hg and Pb using gametophytes of Pityrogramma calomelanos. Int. J. Phytoremediation 2021, 23, 307–315. [Google Scholar] [CrossRef]
- Yang, X.; Chen, H.; Xu, W.; He, Z.; Ma, M. Hyperaccumulation of arsenic by callus, sporophytes and gametophytes of Pteris vittata cultured. Plant Cell Rep. 2007, 26, 1889–1897. [Google Scholar] [CrossRef]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium toxicity in plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.J.M.; Walker, P. Physiological responses of plants to heavy metals and the quantification of tolerance and toxicity. Chem. Speciat. Bioavailab. 1989, 1, 7–17. [Google Scholar] [CrossRef]
- Baker, A.J.M.; McGrath, S.P.; Sidoli, C.M.D.; Reeves, R.D. The possibility of in situ heavy metal decontamination of polluted soils using crops of metal-accumulating plants. Resour. Conserv. Recycl. 1994, 11, 41–49. [Google Scholar] [CrossRef]
- Morel, J.L.; Echevarria, G.; Goncharova, N. Phytoremediation of Metal-Contaminated Soils Czeck Republic, 1st ed.; Springer Science & Business Media: Dordrecht, The Netherlands, 2006; pp. 25–52. [Google Scholar]
- Branquinho, C.; Serrano, H.C.; Pinto, M.J.; Martins-Loução, M.A. Revisiting the plant hyperaccumulation criteria to rare plants and earth abundant elements. Environ. Pollut. 2007, 146, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Choo, T.P.; Lee, C.K.; Low, K.S.; Hishamuddin, O. Accumulation of chromium (VI) from aqueous solutions using water lilies (Nymphaea spontanea). Chemosphere 2006, 62, 961–967. [Google Scholar] [CrossRef]
- Staves, R.P.; Knaus, R.M. Chromium removal from water by three species of dyckweeds. Aquat. Bot. 1985, 23, 261–273. [Google Scholar] [CrossRef]
- Mishra, V.K.; Tripathi, B.D. Accumulation of chromium and zinc from aqueous solutions using water hyacinth (Eichhornia crassipes). J. Hazard. Mater. 2009, 164, 1059–1063. [Google Scholar] [CrossRef]
- Srivastav, R.K.; Gupta, S.K.; Nigam, K.D.P.; Vasudevan, P. Treatment of chromium and nickel in wastewater by using aquatic plants. Water Res. 1994, 28, 1631–1638. [Google Scholar] [CrossRef]
- Namasivayam, C.; Höll, W.H. Chromium (III) removal in tannery waste waters using Chinese Reed (Miscanthus Sinensis), a fast growing plant. Holz Als Roh-Und Werkst. 2004, 62, 74–80. [Google Scholar] [CrossRef]
- Watrud, L.; Maggard, S.; Shiroyama, T.; Coleman, C.; Johnson, M.; Donegan, K.; Di Giovanni, G.; Porteous, L.; Lee, E. Bracken (Pteridium aquilinum L.) frond biomass and rhizosphere microbial community characteristics are correlated to edaphic factors. Plant Soil. 2003, 249, 359–371. [Google Scholar] [CrossRef]
- Riege, D.; Del Moral, R. Differential Tree Colonization of Old Fields in a Temperate Rain Forest. Am. Midl. Nat. 2004, 151, 251–264. [Google Scholar] [CrossRef]
- Gliessman, S. The establishment of bracken following fire in tropical habitats. Am. Fern J. 1978, 68, 41–44. [Google Scholar] [CrossRef]
- Dolling, A. The vegetative spread of Pteridium aquilinum in a hemiboreal forest invasion or revegetation? For. Ecol. Manag. 1999, 24, 177–184. [Google Scholar] [CrossRef]
- Tinklin, R.; Bowling, D. The Water Relations of Bracken: A Preliminary Study. J. Ecol. 1969, 57, 669–671. [Google Scholar] [CrossRef]
- Navar, V.; Fernandez, E.; Del Amo, S. Allelopathic effects of green fronds of Pteridium aquilinum on cultivated plants, weeds, phytopathogenic fungi and bacteria. Agric. Ecosyst. Environ. 1987, 18, 357–379. [Google Scholar] [CrossRef]
- Lawton, J.H.; MacGarvin, M.; Heads, P.A. Effects of altitude on the abundance and species richness of insect herbivores on bracken. J. Anim. Ecol. 1987, 56, 147–160. [Google Scholar] [CrossRef]
Figure 1.
Gametophyte sensitivity to Cr VI. Biomass (A), chlorophyll a (B), chlorophyll b (C). For every treatment, mean ± SEM is presented. * indicates significant differences between treatments and control and in the time interval (p < 0.05); n = 5 and n = 3 for biomass and chlorophyll experiments, respectively.
Figure 1.
Gametophyte sensitivity to Cr VI. Biomass (A), chlorophyll a (B), chlorophyll b (C). For every treatment, mean ± SEM is presented. * indicates significant differences between treatments and control and in the time interval (p < 0.05); n = 5 and n = 3 for biomass and chlorophyll experiments, respectively.

Figure 2.
Cr VI sensitivity of sporophytes. Biomass of aerial parts (A) and underground parts (B), chlorophyll a (C), chlorophyll b (D). For every treatment, mean ± SEM is presented. * indicates significant differences between treatments and control and in the time interval (p < 0.05); n = 5 for biomass and chlorophyll experiments, respectively.
Figure 2.
Cr VI sensitivity of sporophytes. Biomass of aerial parts (A) and underground parts (B), chlorophyll a (C), chlorophyll b (D). For every treatment, mean ± SEM is presented. * indicates significant differences between treatments and control and in the time interval (p < 0.05); n = 5 for biomass and chlorophyll experiments, respectively.

Figure 3.
Absorption and translocation of Cr VI. Cr accumulation in gametophyte (A), Cr content in aerial parts of sporophyte (B), Cr absorption in underground parts of sporophyte (C), translocation index in sporophyte (D). For every treatment, mean ± SEM are presented. * indicates significant differences between treatments and control and in the time interval, p < 0.05, n = 5.
Figure 3.
Absorption and translocation of Cr VI. Cr accumulation in gametophyte (A), Cr content in aerial parts of sporophyte (B), Cr absorption in underground parts of sporophyte (C), translocation index in sporophyte (D). For every treatment, mean ± SEM are presented. * indicates significant differences between treatments and control and in the time interval, p < 0.05, n = 5.

Figure 4.
Remotion percentage of Cr VI from substrate in gametophytes (A) and sporophytes (B). For every treatment, mean ± SEM is presented. * indicates significant differences among treatments vs. control and in the time Interval, p < 0.05, n = 3.
Figure 4.
Remotion percentage of Cr VI from substrate in gametophytes (A) and sporophytes (B). For every treatment, mean ± SEM is presented. * indicates significant differences among treatments vs. control and in the time Interval, p < 0.05, n = 3.

Figure 5.
Proposed mechanism for Cr VI accumulation in plants. (A) Entrance of Cr VI through the roots and/or rhizome, possibly by sulfide transporters. (B) Intracellular (cortex cells) reduction to Cr III by interacting with organic matter. Both trivalent or hexavalent forms are chelated and transported into vacuoles, where the largest amount is deposited. (C) The remaining Cr fraction not accumulated in cortex cells is transported to aerial parts through xylem. (D) The Cr is deposited into mesophile cells in both palisade and spongy tissues. (E) Once inside the cells of photosynthetic tissue, the chelated Cr is deposited into vacuoles, chloroplasts or mitochondria. It is even secreted outside the plant through epidermal cells (CA = chelating agent; Chromium = Cr; transporter= TAR).
Figure 5.
Proposed mechanism for Cr VI accumulation in plants. (A) Entrance of Cr VI through the roots and/or rhizome, possibly by sulfide transporters. (B) Intracellular (cortex cells) reduction to Cr III by interacting with organic matter. Both trivalent or hexavalent forms are chelated and transported into vacuoles, where the largest amount is deposited. (C) The remaining Cr fraction not accumulated in cortex cells is transported to aerial parts through xylem. (D) The Cr is deposited into mesophile cells in both palisade and spongy tissues. (E) Once inside the cells of photosynthetic tissue, the chelated Cr is deposited into vacuoles, chloroplasts or mitochondria. It is even secreted outside the plant through epidermal cells (CA = chelating agent; Chromium = Cr; transporter= TAR).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Eslava-Silva, F.d.J.; Muñíz-Díaz de León, M.E.; Jiménez-Estrada, M. Pteridium aquilinum (Dennstaedtiaceae), a Novel Hyperaccumulator Species of Hexavalent Chromium. Appl. Sci. 2023, 13, 5621. https://doi.org/10.3390/app13095621
AMA Style
Eslava-Silva FdJ, Muñíz-Díaz de León ME, Jiménez-Estrada M. Pteridium aquilinum (Dennstaedtiaceae), a Novel Hyperaccumulator Species of Hexavalent Chromium. Applied Sciences. 2023; 13(9):5621. https://doi.org/10.3390/app13095621
Chicago/Turabian StyleEslava-Silva, Felipe de Jesús, María Eugenia Muñíz-Díaz de León, and Manuel Jiménez-Estrada. 2023. "Pteridium aquilinum (Dennstaedtiaceae), a Novel Hyperaccumulator Species of Hexavalent Chromium" Applied Sciences 13, no. 9: 5621. https://doi.org/10.3390/app13095621
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.