
An Improved Reference Gene for Detection of “Candidatus Liberibacter asiaticus” Associated with Citrus Huanglongbing by qPCR and Digital Droplet PCR Assays

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Analysis of a Single Copy Nuclear Gene in Aurantioideae
2.2. Primers and Probes for Detecting Internal Control Gene Malate Dehydrogenase
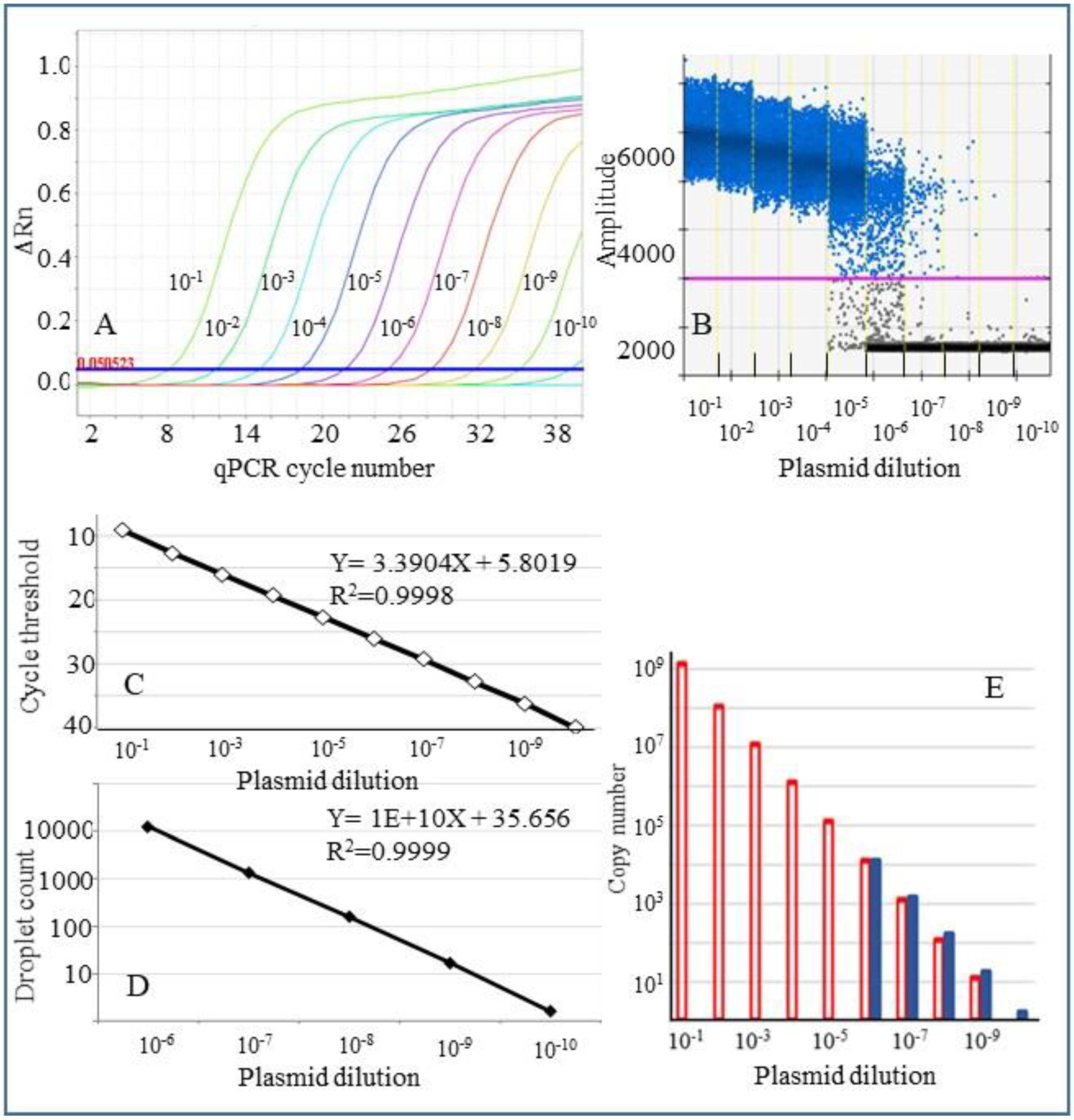
2.3. Comparison of qPCR and ddPCR Assays Using a Plasmid Control for Detection of CLas
2.4. Analysis of Plant Samples Using Duplex qPCR and Singleplex ddPCR Assays
2.5. Analysis of Samples from Diverse Taxa for CLas by qPCR Using COX and MDH as Reference Genes
2.6. Comparison of COX and MDH as Internal Controls for Detection of Two Target Genes of CLas by qPCR
2.7. ddPCR of Reference Gene MDH and CLas in Singleplex and Duplex Assays
3. Discussion
3.1. HLB Situation in California
3.2. Importance of Detecting Samples with Low Titer of CLas
3.3. Importance of Reference Genes
3.4. COX Is Suboptimal as a Reference Gene
3.5. Selection of an Alternate Single Copy Universal Reference Gene
3.6. Reference Gene Requirement in Digital PCR
3.7. Importance of Appropriate Serial Dilutions
4. Materials and Methods
4.1. Sequencing of a Single Copy Nuclear Gene from Members of Aurantioideae
4.2. Design of Primers and Probes for Detecting Internal Control Gene, Malate Dehydrogenase
4.3. Standard Curves for qPCR and ddPCR Assays
4.4. qPCR Assays and Analysis of Results
4.5. Controls for qPCR and ddPCR Assays
4.6. Comparison of COX and MDH as Internal Controls for Detection of CLas by qPCR
4.7. Analysis of Samples from Diverse Taxa for Detection of CLas by qPCR Using COX and MDH as Reference Genes
4.8. Analysis of Citrus Samples by Duplex qPCR and Singleplex ddPCR
4.9. Duplex ddPCR Reactions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bove, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar] [CrossRef]
- da Graca, J.V.; Korsten, L. Citrus huanglongbing: Review, present status and future strategies. In Diseases of Fruits and Vegetables; Naqui, S.A.M.H., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 229–245. [Google Scholar]
- Halbert, S.E.; Manjunath, K.L. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in Florida. Fla. Entomol. 2004, 87, 330–353. [Google Scholar] [CrossRef]
- Kumagai, L.B.; LeVesque, C.S.; Blomquist, C.L.; Madishetty, K.; Guo, Y.; Woods, P.W.; Rooney-Latham, S.; Rascoe, J.; Gallindo, T.; Schnabel, D.; et al. First Report of Candidatus Liberibacter asiaticus Associated with Citrus Huanglongbing in California. Plant Dis. 2013, 97, 283. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Rascoe, J.; Kumagai, L.B.; Keremane, M.L.; Nakhla, M.K. Citrus Huanglongbing (HLB) Discoveries in California in 2015 and 2012 are of Different Genotypes of Candidates Liberibacter asiaticus (cLas) by Double-locus Genomic Variation Analysis. Plant Dis. 2016, 100, 645. [Google Scholar] [CrossRef]
- Manjunath, K.L.; Halbert, S.E.; Ramadugu, C.; Webb, S.; Lee, R.F. Detection of ‘Candidatus Liberibacter asiaticus’ in Diaphorina citri and its importance in the management of Citrus huanglongbing in Florida. Phytopathology 2008, 98, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Li, W.B.; Hartung, J.S.; Levy, L. Quantitative real-time PCR for detection and identification of Candidatus Liberibacter species associated with citrus huanglongbing. J. Microbiol. Meth. 2006, 66, 104–115. [Google Scholar] [CrossRef]
- Lee, J.A.; Halbert, S.E.; Dawson, W.O.; Robertson, C.J.; Keesling, J.E.; Singer, B.H. Asymptomatic spread of huanglongbing and implications for disease control. Proc. Natl. Acad. Sci. USA 2015, 112, 7605–7610. [Google Scholar] [CrossRef] [Green Version]
- Chiyaka, C.; Singer, B.H.; Halbert, S.E.; Morris, J.G.; van Bruggen, A.H.C. Modeling huanglongbing transmission within a citrus tree. Proc. Natl. Acad. Sci. USA 2012, 109, 12213–12218. [Google Scholar] [CrossRef] [Green Version]
- Griffin, K.L.; Anderson, O.R.; Tissue, D.T.; Turnbull, M.H.; Whitehead, D. Variations in dark respiration and mitochondrial numbers within needles of Pinus radiata grown in ambient or elevated CO2 partial pressure. Tree Physiol. 2004, 24, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Thompson, P.; Bowsher, C.G.; Tobin, A.K. Heterogeneity of mitochondrial protein biogenesis during primary leaf development in barley. Plant Physiol. 1998, 118, 1089–1099. [Google Scholar] [CrossRef] [Green Version]
- Kozera, B.; Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 2013, 54, 391–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.; Hu, W.; Xia, Z.; Zhou, X.; Wang, W. Validation of Reference Genes for Relative Quantitative Gene Expression Studies in Cassava (Manihot esculenta Crantz) by Using Quantitative Real-Time PCR. Front. Plant Sci. 2016, 7, 680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hindson, B.J.; Ness, K.D.; Masquelier, D.A.; Belgrader, P.; Heredia, N.J.; Makarewicz, A.J.; Bright, I.J.; Lucero, M.Y.; Hiddessen, A.L.; Legler, T.C.; et al. High-throughput droplet digital PCR system for absolute quantitation of DNA copy number. Anal. Chem. 2011, 83, 8604–8610. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xia, Q.Y.; Yin, Y.P.; Wang, Z.K. Comparison of Droplet Digital PCR and Quantitative PCR Assays for Quantitative Detection of Xanthomonas citri Subsp citri. PLoS ONE 2016, 11, e0159004. [Google Scholar] [CrossRef] [Green Version]
- Maheshwari, Y.; Selvaraj, V.; Hajeri, S.; Yokomi, R. Application of droplet digital PCR for quantitative detection of Spiroplasma citri in comparison with real time PCR. PLoS ONE 2017, 12, e0184751. [Google Scholar] [CrossRef] [Green Version]
- Selvaraj, V.; Maheshwari, Y.; Hajeri, S.; Chen, J.; McCollum, T.G.; Yokomi, R. Development of a duplex droplet digital PCR assay for absolute quantitative detection of ‘Candidatus Liberibacter asiaticus’. PLoS ONE 2018, 13, e0197184. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Liu, X.L.; Lou, B.H.; Zhou, C.Y.; Wang, X.F. Development of a sensitive and reliable droplet digital PCR assay for the detection of ‘Candidatus Liberibacter asiaticus’. J. Integr. Agric. 2018, 17, 483–487. [Google Scholar] [CrossRef] [Green Version]
- Van Dingenen, J.; Blomme, J.; Gonzalez, N.; Inze, D. Plants grow with a little help from their organelle friends. J. Exp. Bot. 2016, 67, 6267–6281. [Google Scholar] [CrossRef]
- Takanashi, H.; Ohnishi, T.; Mogi, M.; Okamoto, T.; Arimura, S.; Tsutsumi, N. Studies of mitochondrial morphology and DNA amount in the rice egg cell. Curr. Genet. 2010, 56, 33–41. [Google Scholar] [CrossRef]
- Whale, A.S.; Huggett, J.F.; Tzonev, S. Fundamentals of multiplexing with digital PCR. Biomol. Detect. Quantif. 2016, 10, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Ramadugu, C.; Pfeil, B.E.; Keremane, M.L.; Lee, R.F.; Maureira-Butler, I.J.; Roose, M.L. A six nuclear gene phylogeny of Citrus (Rutaceae) taking into account hybridization and lineage sorting. PLoS ONE 2013, 8, e68410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J.; Clustal, W. Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; p. 352. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Jagoueix, S.; Bove, J.M.; Garnier, M. PCR detection of the two ‘Candidatus’ Liberobacter species associated with greening disease of citrus. Mol. Cell. Probe 1996, 10, 43–50. [Google Scholar] [CrossRef]
- Gallup, J.M.; Ackermann, M.R. The ‘PREXCEL-Q Method’ for qPCR. Int. J. Biomed. Sci. 2008, 4, 273–293. [Google Scholar] [PubMed]
- Ramadugu, C.; Keremane, M.L.; Halbert, S.E.; Duan, Y.P.; Roose, M.L.; Stover, E.; Lee, R.F. Long-Term Field Evaluation Reveals Huanglongbing Resistance in Citrus Relatives. Plant Dis. 2016, 100, 1858–1869. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Xu, M.R.; Bao, M.L.; Wu, F.N.; Chen, J.C.; Deng, X.L. Unusual Five Copies and Dual Forms of nrdB in ‘Candidatus Liberibacter asiaticus’: Biological Implications and PCR Detection Application. Sci. Rep. 2016, 6, 39020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coletta, H.D.; Takita, M.A.; Targon, M.L.P.N.; Machado, M.A. Analysis of 16S rDNA sequences from citrus huanglongbing bacteria reveal a different ‘Ca. Liberibacter’ strain associated with citrus disease in Sao Paulo. Plant Dis. 2005, 89, 848–852. [Google Scholar] [CrossRef]
- Teixeira, D.D.C.; Saillard, C.; Eveillard, S.; Danet, J.L.; Costa, P.I.D.; Ayres, A.J.; Bove, J. ‘Candidatus Liberibacter americanus’, associated with citrus huanglongbing (greening disease) in Sao Paulo State, Brazil. Int. J. Syst. Evol. Microbiol. 2005, 55, 1857–1862. [Google Scholar] [CrossRef] [Green Version]
- Halbert, S.E. The discovery of huanglongbing in Florida. In Proceedings of the 2nd International Citrus Canker and Huanglongbing Research Workshop, Orlando, FL, USA, 7–11 November 2005; p. H3. [Google Scholar]
- Bartels, D.; Cook, G. Update on the hot spot cluster analysis of Ct-values from Asian citrus psyllid samples. J. Citrus Pathol. 2017, 4, 3–4. [Google Scholar]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol. 2000, 25, 169–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Huggett, J.F.; Foy, C.A.; Benes, V.; Emslie, K.; Garson, J.A.; Haynes, R.; Hellemans, J.; Kubista, M.; Mueller, R.D.; Nolan, T.; et al. The digital MIQE guidelines: Minimum Information for Publication of Quantitative Digital PCR Experiments. Clin. Chem. 2013, 59, 892–902. [Google Scholar] [CrossRef] [PubMed]
- Westbrook, C.J.; Hall, D.G.; Stover, E.; Duan, Y.P.; Lee, R.F. Colonization of Citrus and Citrus-related Germplasm by Diaphorina citri (Hemiptera: Psyllidae). Hortscience 2011, 46, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Halbert, S.E.; Keremane, M.L.; Ramadugu, C.; Lee, R.F. Incidence of huanglongbing-associated ‘Candidatus Liberibacter asiaticus’ in Diaphorina citri (Hemiptera: Psyllidae) collected from plants for sale in Florida. Fla. Entomol. 2012, 95, 620–627. [Google Scholar] [CrossRef] [Green Version]
- Kralik, P.; Ricchi, M. A Basic Guide to Real Time PCR in Microbial Diagnostics: Definitions, Parameters, and Everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.; Zhao, Z.; Qian, C.; Sui, Y.; Malik, A.A.; Chen, J. Selection of appropriate reference genes for gene expression studies by quantitative real-time polymerase chain reaction in cucumber. Anal. Biochem. 2010, 399, 257–261. [Google Scholar] [CrossRef] [PubMed]
- LeVesque, C.; McRoberts, N. Comparative study of early detection techniques: Texas 2 study. Citrograph 2017, 8, 44–47. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ to 3′) | Target | Reference |
|---|---|---|---|
| CIT205a | GCGCGTATGCgAATACGAGCGGCA | CLas 16s forward (OA1) * | [26] |
| CIT206 | GCCTCGCGACTTCGCAACCCAT | CLas 16s reverse (OIc) | |
| CIT295a | TCGAGCGCGTATGCgAATACG | Clas-16s forward * | [7] |
| CIT298 | TGCGTTATCCCGTAGAAAAAGGTAG | Clas-16s reverse | |
| CIT301 | CAGACGGGTGAGTAACGCG | Clas-16s probe | |
| CIT315 | GTATGCCACGTCGCATTCCAGA | COX-F forward | [7] |
| CIT316 | ATCCAGATGCTTACGCTGG | COX-P probe ** | |
| CIT317 | GCCAAAACTGCTAAGGGCATTC | COX-R reverse | |
| CIT637 | GCTCCTGTGGAAGAGACCC | MDH forward | This study |
| CIT638 | GCTCCAGAGATGACCAAAC | MDH reverse | |
| CIT1249 | TTGCATGCAGGGACTGATAG | MDH forward | This study |
| CIT1250 | GATTGTTGACAGGCTCATGTTC | MDH reverse | |
| CIT1251 | ACACTTCAAGAAGCCTTGGGCTCA | MDH Probe ** |
| CRC# | Genus | Species | Variety | COX | MDH | Difference |
|---|---|---|---|---|---|---|
| 3564 | Citrus | lycopersicaformis | Kokni Orange | 19.05 | 27.17 | 8.12 |
| 3546 | Citrus | medica | South Coast Field Station | 18.26 | 26.57 | 8.31 |
| 3673 | Microcitrus | australis | Australian round lime | 17.77 | 26.19 | 8.42 |
| 1482 | Citrus | limettioides | Palestine sweet lime | 17.95 | 26.65 | 8.70 |
| 3474 | Citrus | intermedia | Yama-mikan sour orange | 17.98 | 26.79 | 8.81 |
| 3149 | Citrus | benikoji | Unnamed tangor | 19.53 | 28.42 | 8.89 |
| 3514 | Balsamocitrus | daweii | Uganda powder flask | 18.52 | 27.58 | 9.06 |
| 3052 | Citrus | latipes | Khasi papeda | 18.56 | 27.93 | 9.37 |
| 4107 | Severinia | buxifolia | Chinese box orange | 18.49 | 27.94 | 9.45 |
| 1484 | Microcitrus | australasica | Australian finger lime | 18.70 | 28.25 | 9.54 |
| 3225 | Citrus | maderaspatana | Kitchli Sour orange hybrid | 18.62 | 28.24 | 9.62 |
| 1455 | Citrus | webberi | Kalpi papeda | 18.28 | 28.05 | 9.77 |
| 3752 | Citrus | reticulata | Som Keowan mandarin | 16.56 | 26.40 | 9.84 |
| 1485 | Microcitrus | hybrid | Sydney Hybrid | 20.47 | 30.32 | 9.85 |
| 3771 | X Citroncirus | sp. | Swingle citrumelo | 17.79 | 27.78 | 9.99 |
| 4105 | Eremocitrus | glauca | Australian Desert lime | 18.82 | 28.83 | 10.01 |
| 2588 | Citrus | taiwanica | Nansho daidai sour orange | 18.06 | 28.08 | 10.02 |
| 3959 | Citrus | maxima | Egami Buntan pummelo | 18.68 | 28.75 | 10.08 |
| 3842 | Citrus | celebica | Alemow hybrid | 17.95 | 28.14 | 10.19 |
| 3957 | X Citroncirus | sp. | X639 trifoliate hybrid | 16.83 | 27.04 | 10.20 |
| 2320 | Citrus | longispina | Winged lime | 18.64 | 28.90 | 10.26 |
| 2317 | Citrus | excelsa | Limon Real papeda | 19.37 | 29.64 | 10.26 |
| 2485 | Citrus | amblycarpa | Nasnaran mandarin | 17.56 | 27.89 | 10.33 |
| 3147 | Citrus | leiocarpa | Koji mandarin | 19.55 | 29.98 | 10.42 |
| 3907 | Citrus | hassaku | Hassaku pummelo hybrid | 17.84 | 28.27 | 10.43 |
| 1491 | Severinia | buxifolia | Chinese box orange | 19.13 | 29.80 | 10.67 |
| 2427 | Citrus | davaoensis | Davao lemon | 16.28 | 26.99 | 10.71 |
| 3549 | Poncirus | trifoliata | Simmons trifoliate | 18.72 | 29.60 | 10.89 |
| 4007 | Poncirus | trifoliata | Little-Leaf trifoliate | 16.68 | 27.62 | 10.93 |
| 2879 | Hesperethusa | crenulata | Hesperethusa | 18.87 | 29.83 | 10.96 |
| 301 | X Citroncirus | sp. | Rusk citrange | 17.60 | 28.91 | 11.31 |
| 3285 | Glycosmis | pentaphylla | Orange berry | 24.98 | 36.51 | 11.53 |
| 2878 | Aeglopsis | chevalieri | Chevalier’s Aeglopsis | 19.98 | 31.92 | 11.95 |
| 3165 | Bergera | koenigii | Curry leaf | 18.89 | 31.23 | 12.34 |
| 1637 | Murraya | paniculata | Orange Jessamine | 18.30 | 33.74 | 15.44 |
| 3171 | Murraya | paniculata | Hawaiian Mock Orange | 16.94 | 32.70 | 15.76 |
| Dilution | COX Ct | MDH Ct |
|---|---|---|
| 10−1 | 22.27 | 28.81 |
| 10−2 | 25.74 | 32.97 |
| 10−3 | 29.36 | 36.97 |
| 10−4 | 33.49 | 0 |
| 10−5 | 36.38 | 0 |
| 10−6 | 0 | 0 |
| NTC | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keremane, M.L.; McCollum, T.G.; Roose, M.L.; Lee, R.F.; Ramadugu, C. An Improved Reference Gene for Detection of “Candidatus Liberibacter asiaticus” Associated with Citrus Huanglongbing by qPCR and Digital Droplet PCR Assays. Plants 2021, 10, 2111. https://doi.org/10.3390/plants10102111
Keremane ML, McCollum TG, Roose ML, Lee RF, Ramadugu C. An Improved Reference Gene for Detection of “Candidatus Liberibacter asiaticus” Associated with Citrus Huanglongbing by qPCR and Digital Droplet PCR Assays. Plants. 2021; 10(10):2111. https://doi.org/10.3390/plants10102111
Chicago/Turabian StyleKeremane, Manjunath L., Thomas G. McCollum, Mikeal L. Roose, Richard F. Lee, and Chandrika Ramadugu. 2021. "An Improved Reference Gene for Detection of “Candidatus Liberibacter asiaticus” Associated with Citrus Huanglongbing by qPCR and Digital Droplet PCR Assays" Plants 10, no. 10: 2111. https://doi.org/10.3390/plants10102111