
Smilax weniae, a New Species of Smilacaceae from Limestone Areas Bordering Guizhou and Guangxi, China

 and
and
Abstract
:1. Introduction
2. Results
2.1. Morphological Comparison
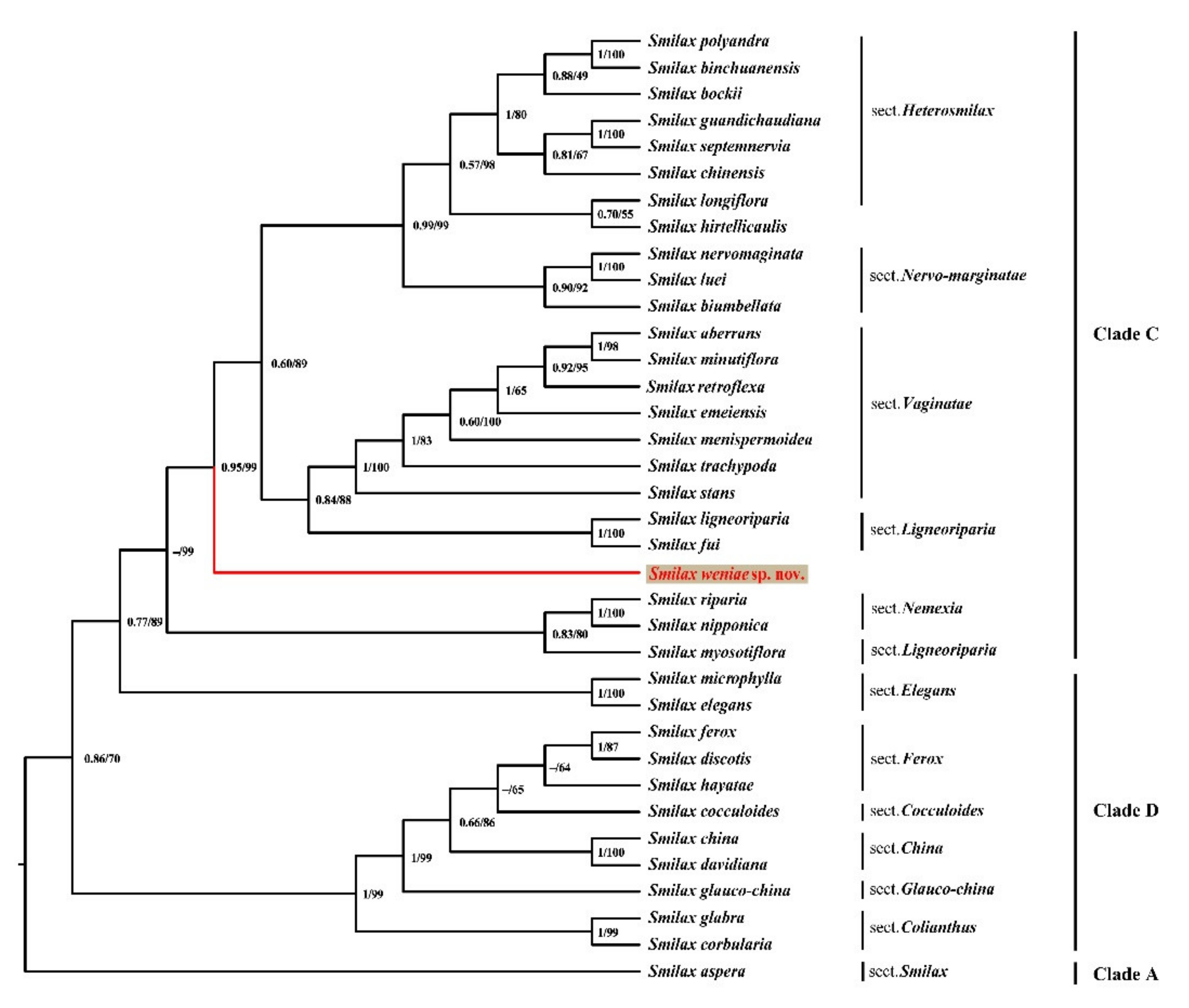
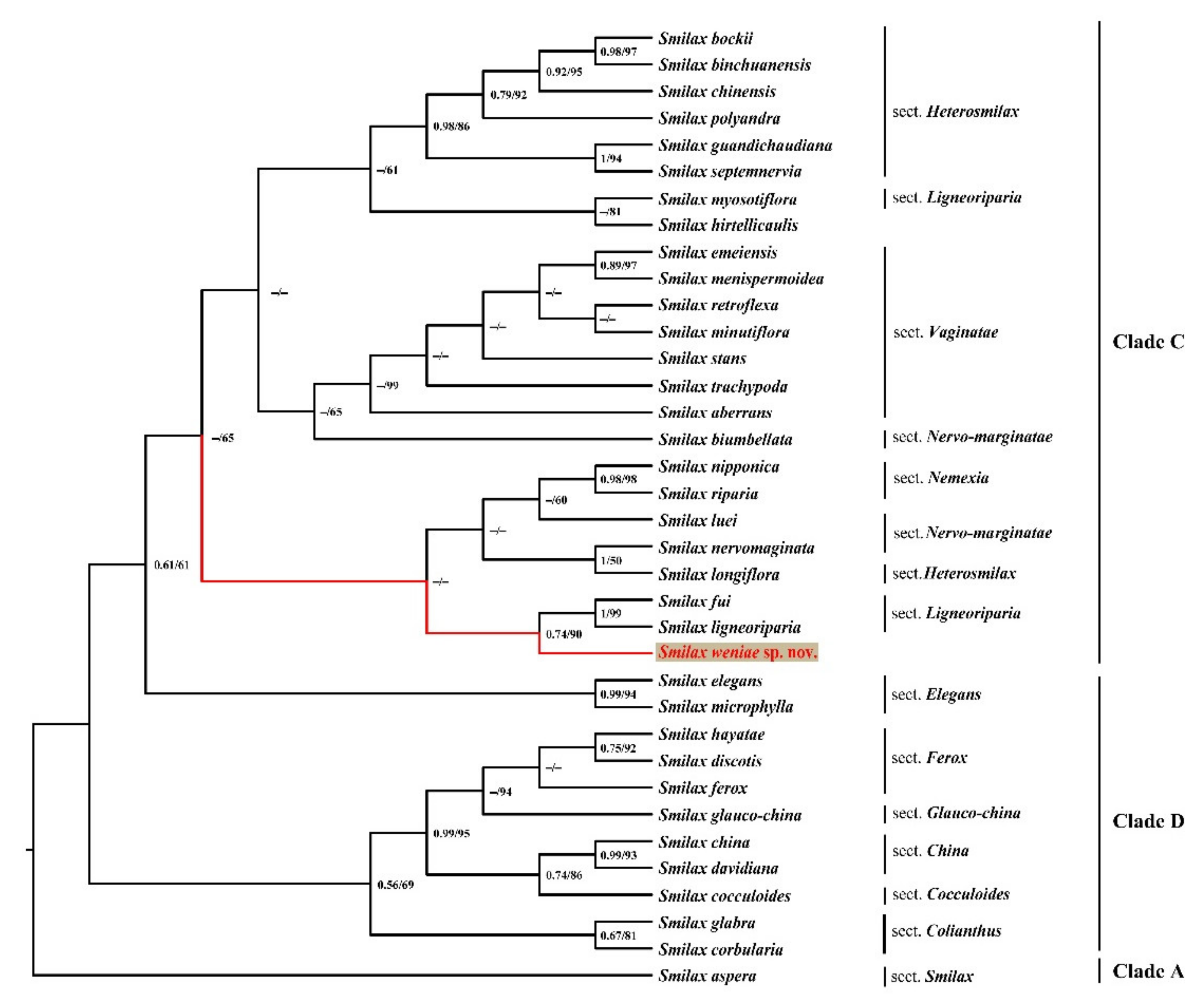
2.2. Molecular Phylogenetic Analyses
2.3. Taxonomic Treatment of the New Species
2.3.1. Diagnosis
2.3.2. Additional Specimens Seen (Paratypes)
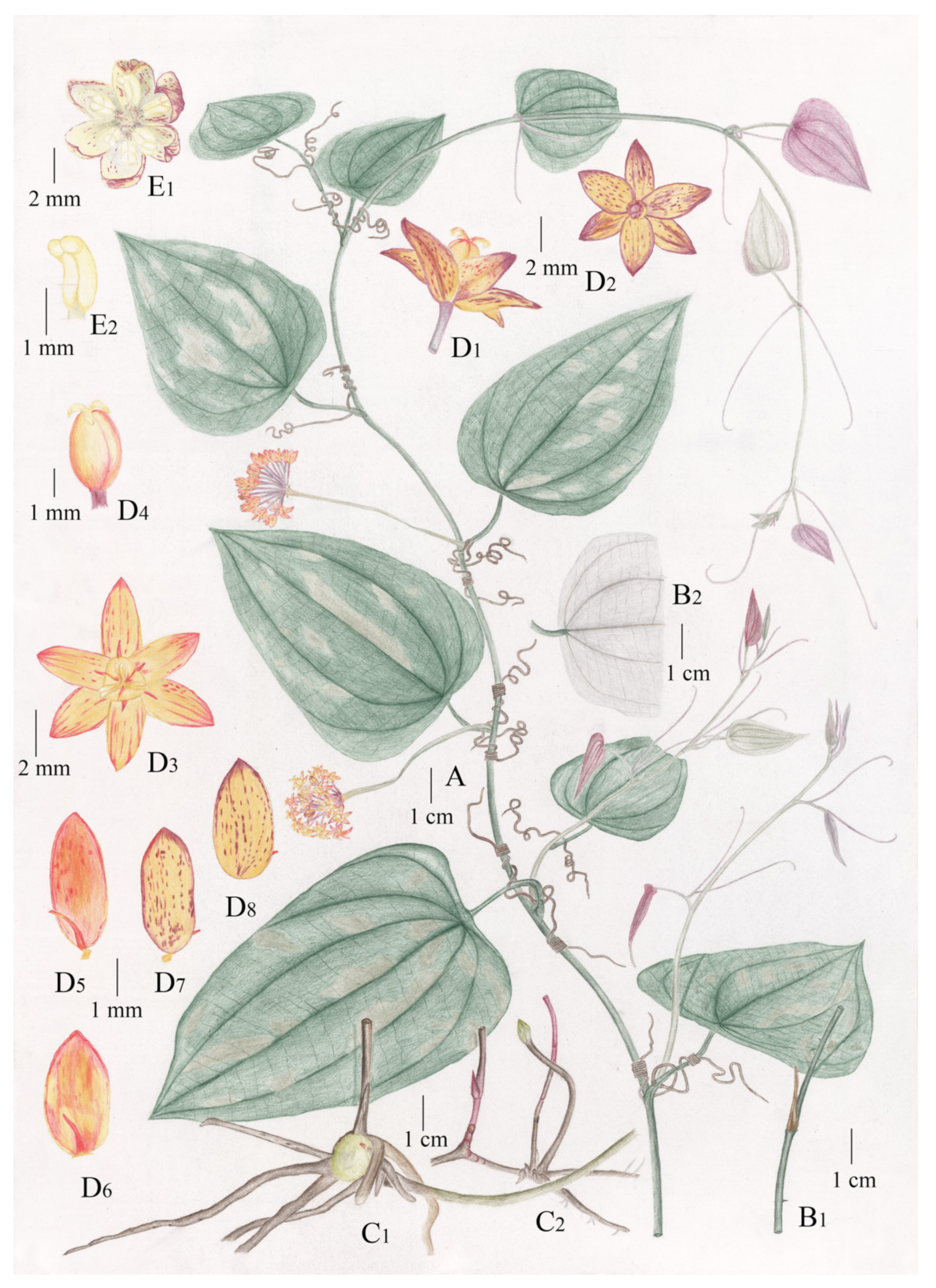
2.3.3. Description
2.3.4. Etymology
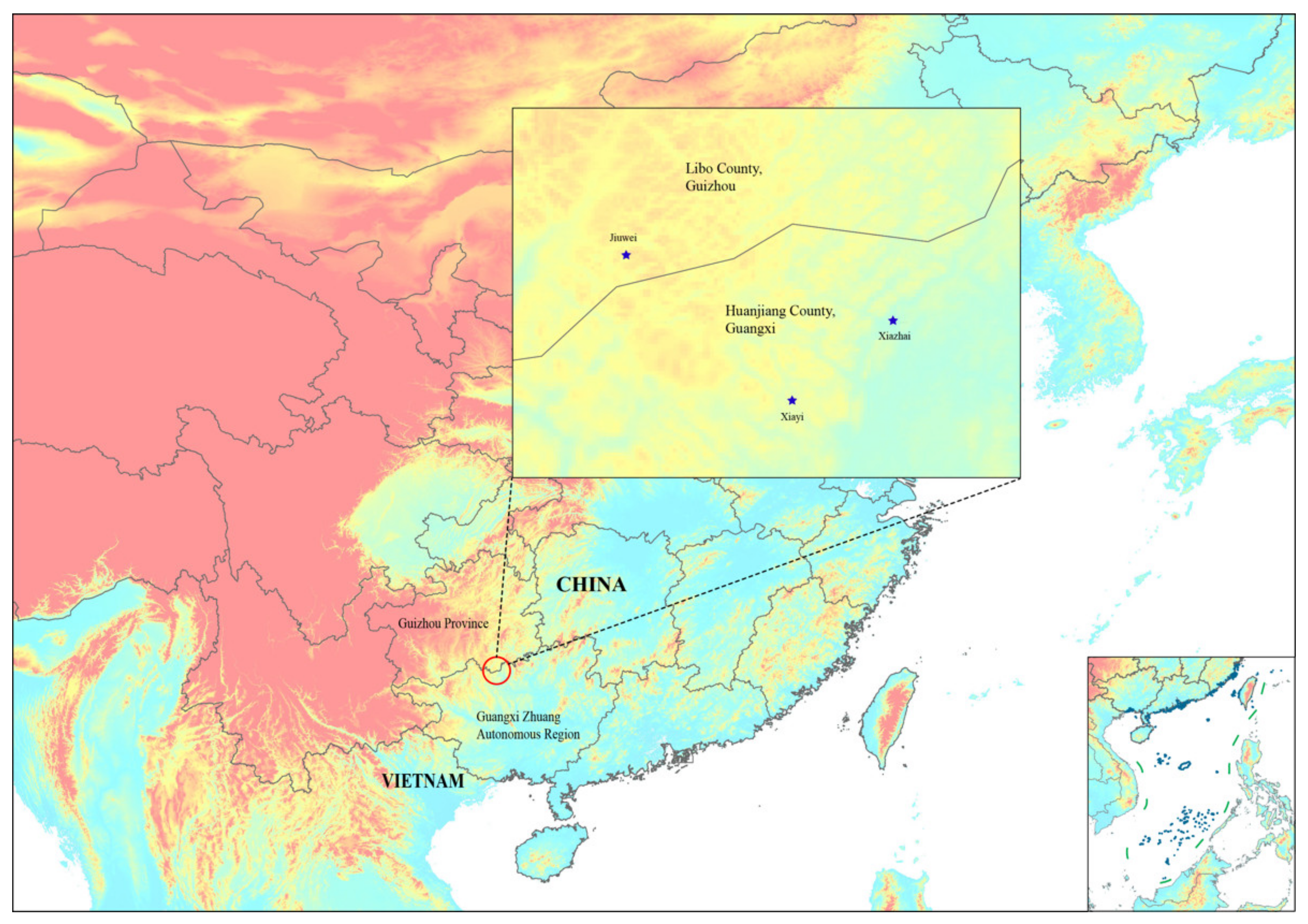
2.3.5. Distribution and Habitat
2.3.6. Conservation Status
2.3.7. Taxonomic and Evolutionary Relationships
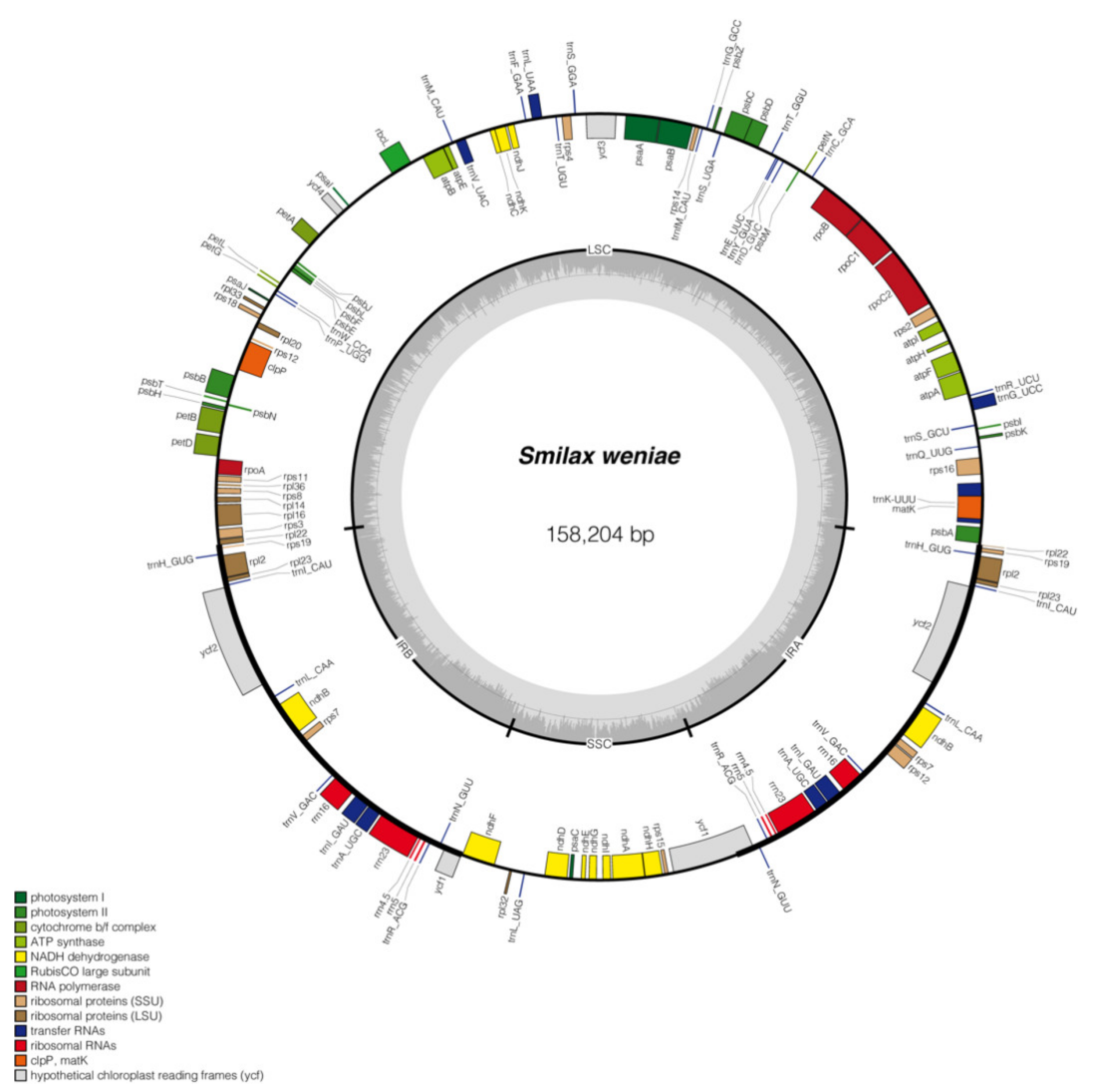
2.4. Characteristics of Plastome
3. Discussion
4. Materials and Methods
4.1. Morphological Observation
4.2. Molecular Methods and Phylogenetic Analyses
4.3. Plastome Sequencing and Analysis of Smilax weniae
4.4. Chromosome Counts
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qi, Z.C.; Cameron, K.M.; Li, P.; Zhao, Y.P.; Shen, C.; Chen, G.C.; Fu, C.X. Phylogenetics, character evolution, and distribution patterns of the greenbriers, Smilacaceae (Liliales), a near-cosmopolitan family of monocots. Bot. J. Linn. Soc. 2013, 173, 535–548. [Google Scholar] [CrossRef] [Green Version]
- Cameron, K.M.; Fu, C.X. A nuclear rDNA phylogeny of Smilax (Smilacaceae). Aliso 2006, 22, 598–605. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.Q.; Koyama, T. Flora of China; Wu, Z.-Y., Raven, P.H., Eds.; Science Press: Beijing, China; Beijing & Missouri Botanical Garden Press: St. Lours, MI, USA, 2000; Volume 24, pp. 96–115. [Google Scholar]
- Koyama, T. A taxonomic revision of the genus Heterosmilax (Smilacaceae). Brittonia 1984, 36, 184–205. [Google Scholar] [CrossRef]
- Ferrufino-Acosta, L. Taxonomic revision of the genus Smilax (Smilacaceae) in Central America and the Caribbean Islands. Willdenowia 2010, 40, 227–280. [Google Scholar] [CrossRef] [Green Version]
- IUCN. Guidelines for Using the IUCN Red List Categories and Criteria. Version 15. Prepared by the Standards and Petitions Committee. 2022. Available online: https://www.iucnredlist.org/resources/redlistguidelines (accessed on 19 February 2022).
- Wagensommer, R.P.; Bartolucci, F.; Fiorentino, M.; Licht, W.; Peccenini, S.; Perrino, E.V.; Venanzoni, R. First record for the flora of Italy and lectotypification of the name Linum elegans (Linaceae). Phytotaxa 2017, 296, 161–170. [Google Scholar] [CrossRef]
- Wagensommer, R.P.; Venanzoni, R. Geranium lucarinii sp. nov. and re-evaluation of G. kikianum (Geraniaceae). Phytotaxa 2021, 489, 252–262. [Google Scholar] [CrossRef]
- Blasco, F.A.; Rubite, R.R.; Cortes, J.C.; Alejandro, G.J.D. Begonia lanuzaensis (sect. Petermannia, Begoniaceae) a new species from Surigao del Sur, Mindanao Island, Philippines. Phytotaxa 2021, 523, 203–207. [Google Scholar] [CrossRef]
- Swanepoel, W.; de Cauwer, V.; Van Wyk, A.E. A new rheophytic species of Syzygium (Myrtaceae) from the lower Kunene River of Angola and Namibia. Phytotaxa 2021, 491, 281–290. [Google Scholar] [CrossRef]
- Chinchilla, I.F. A new tree species of Cupania (Sapindoideae, Sapindaceae) from Quepos, Costa Rica. Phytotaxa 2020, 475, 178–186. [Google Scholar] [CrossRef]
- Kong, H.H.; Wang, A.L.; Li, C.Q.; Fu, C.X. Studies of systematic evolution and karyotypic variation in Smilax and Heterosmilax (Smilacaceae). Acta Phytotaxon. Sin. 2007, 45, 257–273. [Google Scholar] [CrossRef]
- Jin, X.J.; Chen, Y.; Lee, J.; Qi, Z.C.; Liu, L.X.; Li, P.; Fu, C.X. A new species of Smilax (Smilacaceae) from Yunnan, China. Phytotaxa 2016, 275, 159–167. [Google Scholar] [CrossRef]
- Koyama, T. Two New Species of Smilax (Smilacaceae) from Taiwan. Taiwania 1975, 20, 117–122. [Google Scholar] [CrossRef]
- Liu, G.T.; Tsai, K.L.; Hsieh, S.I. New distribution information for Smilax luei T. Koyama (Smilacaceae), a rare plant in Taiwan. Nat. Conserv. Q 2017, 98, 74–80. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 30 August 2021).
- Li, J.L.; Wang, S.; Yu, J.; Wang, L.; Zhou, S.L. A Modified CTAB Protocol for Plant DNA Extraction. Chin. Bull. Bot. 2013, 48, 72–78. [Google Scholar] [CrossRef]
- Fu, C.X.; Kong, H.H.; Qiu, Y.X.; Cameron, K.M. Molecular phylogeny of the East Asian-North American disjunct Smilax sect. Nemexia (Smilacaceae). Int. J. Plant Sci. 2005, 166, 301–309. [Google Scholar] [CrossRef]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetics studies in angiosperms: The Tortoise and The Hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.A.; Drummond, A.J. Tracer v1.6. 2014. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 30 August 2021).
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v. 1.4.3. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 30 August 2021).
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; Claude, W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—versatile and accurate annotation of organelle genomes. Nucleic Acids. Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | S. weniae | S. luei |
|---|---|---|
| Stem | prickless or occasionally prickly | prickless |
| Leaf blade | ovate, peltate, leaf base truncate, retuse or round, 5.5–14.5 cm long, 3.5–9.5 cm wide | lanceolate, peltate, leaf base retuse or round, 3–13 cm long, 1–3.5 cm wide |
| Male flower | tepals connate ca. 1/5, stamens 6 | tepals separated completely, stamens 9 (rarely 10), |
| Female flower | tepals glabrous on both surfaces | tepals minutely verruculose on the outer surface |
| Peduncle | 2.5–7.2(–10) cm long | 1.5–2.5 cm long |
| Family | Species | Voucher | ITS | matK | rbcL | ndhA | ndhF | rpl16 |
|---|---|---|---|---|---|---|---|---|
| Smilacaceae | Smilax weniae P. Li, Z.C. Qi & Yan Liu. sp. nov. | P. Li 009119 (HZU) | OL677459 | OL504525 | OL504528 | OL504526 | OL504527 | OL504529 |
| S. aberrans Gagnep. | C. Fu 20037 (HZU) | JF461346 | JF461377 | MT105074 | KF818408 | KF818438 | JF461422 | |
| S. aspera L. | BQ 0908304 (HZU) | JF461347 | JF461378 | MT105075 | KF818409 | KF818439 | KC511399 | |
| S. biumbellata T. Koyama | C. Fu 20022 (HZU) | JF461351 | JF461383 | MT105084 | MT104985 | MT104889 | JF461427 | |
| S. china L. | C. Fu 20006 (HZU) | JF978671 | JF956367 | JF944328 | KC522265 | KC5213150 | KC511414 | |
| S. cocculoides Warb. ex Diels | C. Fu 0903122-1 (HZU) | JF978680 | JF956376 | JF944337 | KF818413 | KF818443 | KC511416 | |
| S. corbularia Kunth | J. Li 0024298 (KRIBB) | KC511496 | KC511356 | MT105091 | MT104994 | MT104898 | KC511417 | |
| S. davidiana A. DC. | C. Fu Fw 108 (HZU) | KC511498 | KC511358 | MT105093 | KF818414 | KF818444 | KC511421 | |
| S. discotis Warb. | C. Fu Fw 111 (HZU) | JF956388 | KC511359 | JF944349 | KF818415 | KF818445 | KC511422 | |
| S. elegans Wall. | P. Li 0908300-1 (HZU) | JF978692 | JF956392 | JF944353 | KC522275 | KC5213160 | KC511423 | |
| S. emeiensis J.M. Xu | 0610004 (HZU) | JF461353 | JF461385 | MT105097 | MT105000 | MT104904 | JF461429 | |
| S. ferox Wall. ex Kunth | C. Fu 20059 (HZU) | JF978696 | JF956396 | JF944357 | KF818417 | KF818447 | KC511426 | |
| S. fui Z.C. Qi & P. Li | C. Fu 10043 (HZU) | MT104825 | MT104854 | MT105150 | MT105047 | MT104951 | MT105173 | |
| S. glabra Roxb. | C. Fu 903107 (HZU) | JF978699 | JF956399 | JF944360 | KF818418 | KF818448 | KC511427 | |
| S. glauco-china Warb. ex Diels | C. Fu Fw114 (HZU) | JF978706 | JF956406 | JF944367 | KF818420 | KF818450 | KC511429 | |
| S. hayatae Koyama | C. Fu 0903151 (HZU) | MT104828 | MT104857 | MT105103 | KF818422 | KF818452 | MT105176 | |
| S. hirtellicaulis C.Y. Wu & C. Chen ex P. Li. | XJJin.HK01 (HZU) | KX712229 | KX712230 | KX712231 | - | - | - | |
| S. ligneoriparia C.X. Fu & P. Li | P. Li 0904082 (HZU) | JF461359 | JF461395 | MT105118 | MT105015 | MT104919 | JF461438 | |
| S. luei T.Koyama | X. Liu 0809016 (HZU) | KC511503 | JF461396 | MT105120 | MT105017 | MT104921 | KC511440 | |
| S. microphylla C.H. Wright | C. Fu 20007 (HZU) | JF978747 | JF956443 | JF944407 | KC522308 | KC5213193 | KC511445 | |
| S. menispermoidea A. DC. | L. Gao 08898 (KUN) | JF461360 | JF461397 | MT105121 | MT105020 | MT104924 | JF461439 | |
| S. myosotiflora A. DC. | C. Fu 09013 (HZU) | KC511505 | KC511366 | MT105126 | MT105024 | MT104928 | KC511446 | |
| S. nervo-marginata Hayata | C. Fu 2010422 (HZU) | JF461363 | JF461400 | MT105130 | KF818424 | KF818454 | KC511451 | |
| S. nipponica Miq. | C. Fu 950174 (HZU) | AY775244 | JF461402 | MT105132 | KF818425 | KF818455 | JF461443 | |
| S. riparia A. DC. | C. Fu 912691 (HZU) | AY775234 | JF461407 | MT105142 | KC522335 | KC5213220 | AY775217 | |
| S. retroflexa (F.T. Wang & Tang) S.C. Chen | P. Li 0904056 (HZU) | JF461366 | JF461406 | MT105141 | MT105041 | MT104945 | JF461447 | |
| S. stans Maxim. | Y. Wan 0811021 (HZU) | JF461368 | JF461413 | MT105157 | MT105052 | MT104956 | JF461449 | |
| S. minutiflora F.T. Wang | C. Fu 20013 (HZU) | AY775256 | JF461416 | MT105163 | MT105057 | MT104961 | AY775229 | |
| S. trachypoda J.B. Norton | P. Li 0907257 (HZU) | JF461370 | JF461415 | MT105162 | MT105055 | MT104959 | JF461451 | |
| S. chinensis (F.T. Wang) P. Li & C.X. Fu | C. Fu20011 (HZU) | JF461342 | JF461372 | MT105064 | KF818400 | KF818430 | JF461418 | |
| S. gaudichaudiana Kunth. | BQ 0902015 (HZU) | KX394645 | KX432982 | KX394669 | MT104873 | MT104968 | JF461419 | |
| S. bockii Warb. ex Diels | C. Fu 010807 (HZU) | AY775257 | JF461374 | MT105066 | KF818401 | KF818431 | AY775230 | |
| S. longiflora (K.Y. Guan & Noltie) P. Li & C.X. Fu | C. Fu 9908 (HZU) | JF461344 | JF461375 | MT105067 | MT104969 | MT104874 | JF461420 | |
| S. polyandra (F. Gagnep.) P. Li & C.X. Fu | P. Li 0905145 (HZU) | KC511480 | KC511340 | MT105068 | MT104970 | MT104875 | KC511389 | |
| S. septemnervia (F.T. Wang & Tang) P. Li & C.X. Fu | P. Li 0908310 (HZU) | MT104844 | MT104867 | MT105148 | MT104971 | MT104876 | MT105180 | |
| S. binchuanensis P. Li & C.X. Fu | C. Fu 20019 (HZU) | JF976617 | JF461376 | JF941922 | MT104877 | MT104972 | JF461451 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.-Y.; Jin, X.-J.; Zhang, S.-L.; Yang, J.-W.; Fei, S.-P.; Huang, Y.-S.; Liu, Y.; Qi, Z.-C.; Li, P. Smilax weniae, a New Species of Smilacaceae from Limestone Areas Bordering Guizhou and Guangxi, China. Plants 2022, 11, 1032. https://doi.org/10.3390/plants11081032
Feng J-Y, Jin X-J, Zhang S-L, Yang J-W, Fei S-P, Huang Y-S, Liu Y, Qi Z-C, Li P. Smilax weniae, a New Species of Smilacaceae from Limestone Areas Bordering Guizhou and Guangxi, China. Plants. 2022; 11(8):1032. https://doi.org/10.3390/plants11081032
Chicago/Turabian StyleFeng, Jie-Ying, Xin-Jie Jin, Sheng-Lu Zhang, Jia-Wen Yang, Shi-Peng Fei, Yu-Song Huang, Yan Liu, Zhe-Chen Qi, and Pan Li. 2022. "Smilax weniae, a New Species of Smilacaceae from Limestone Areas Bordering Guizhou and Guangxi, China" Plants 11, no. 8: 1032. https://doi.org/10.3390/plants11081032