
Genomic Evaluation of Coffea arabica and Its Wild Relative Coffea racemosa in Mozambique: Settling Resilience Keys for the Coffee Crop in the Context of Climate Change
 , , ,
, , ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
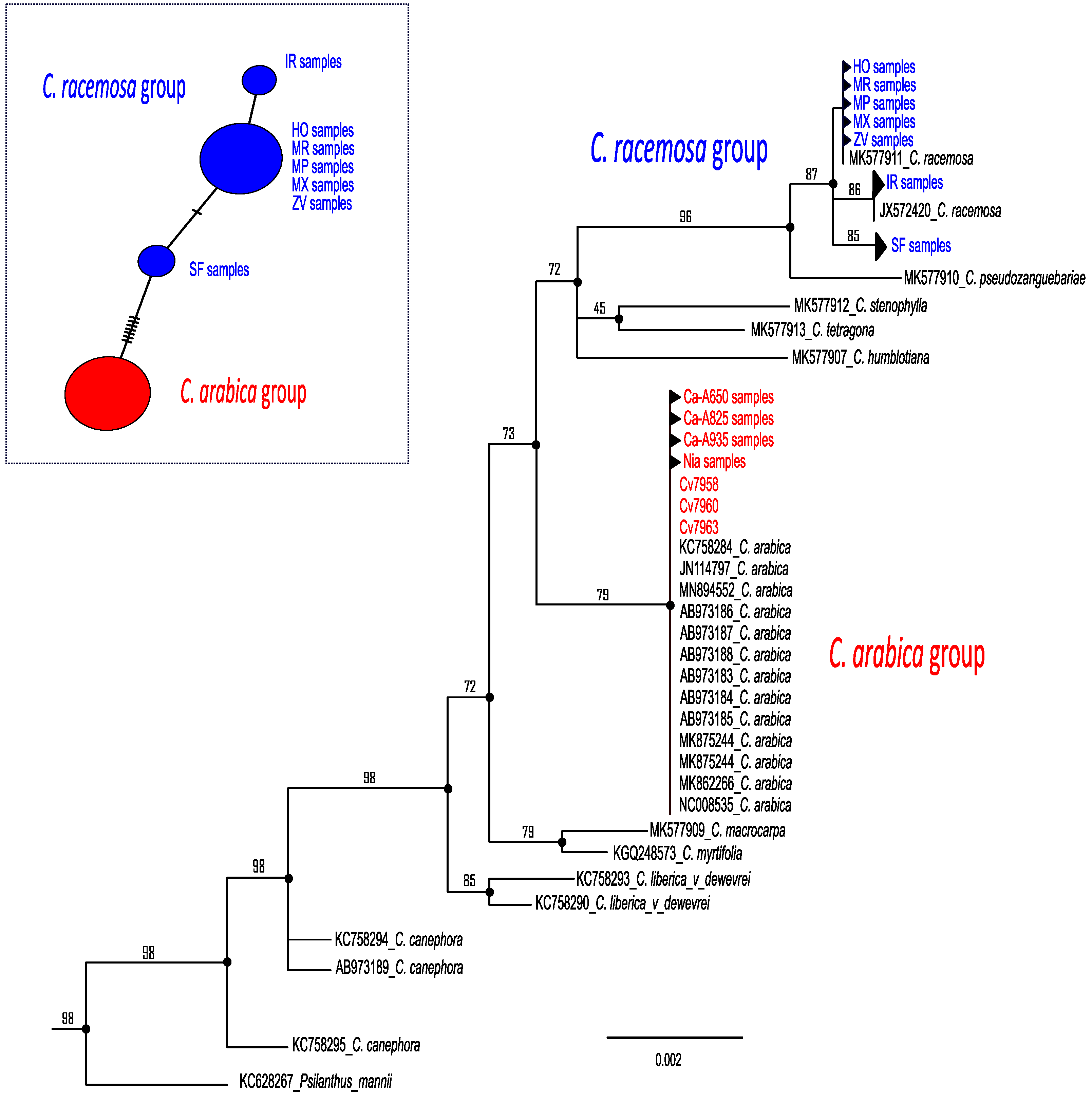
2.1. Plastid Relationships among Coffea Samples
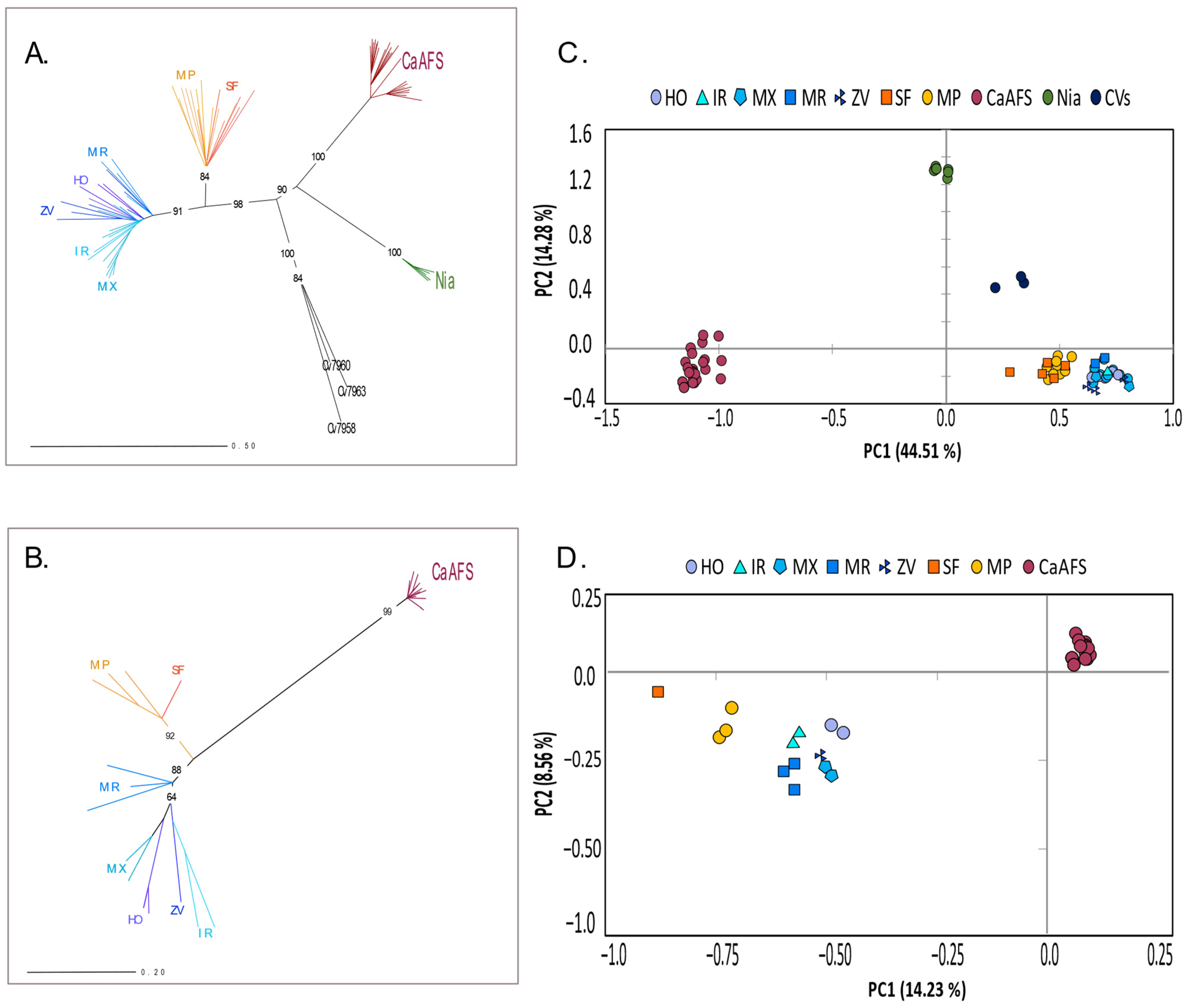
2.2. Genetic Diversity in C. arabica and the Wild Relative C. racemosa
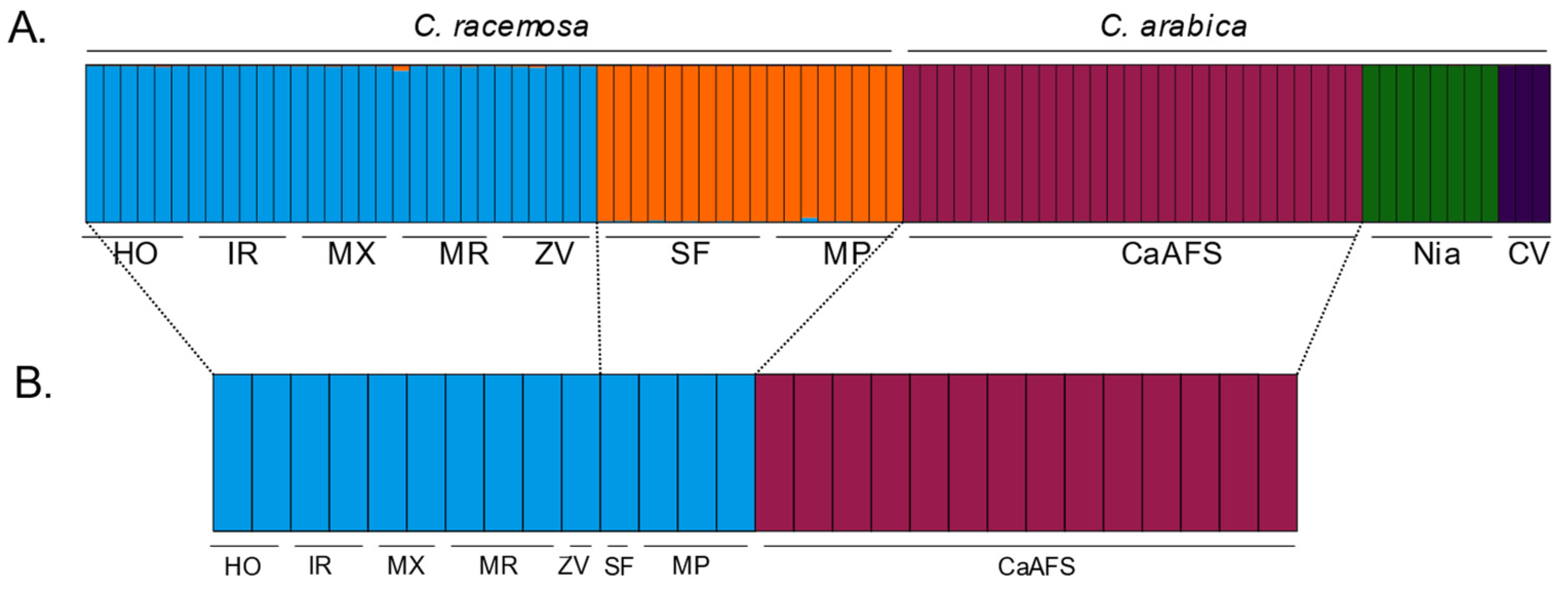
2.3. Genetic Structure
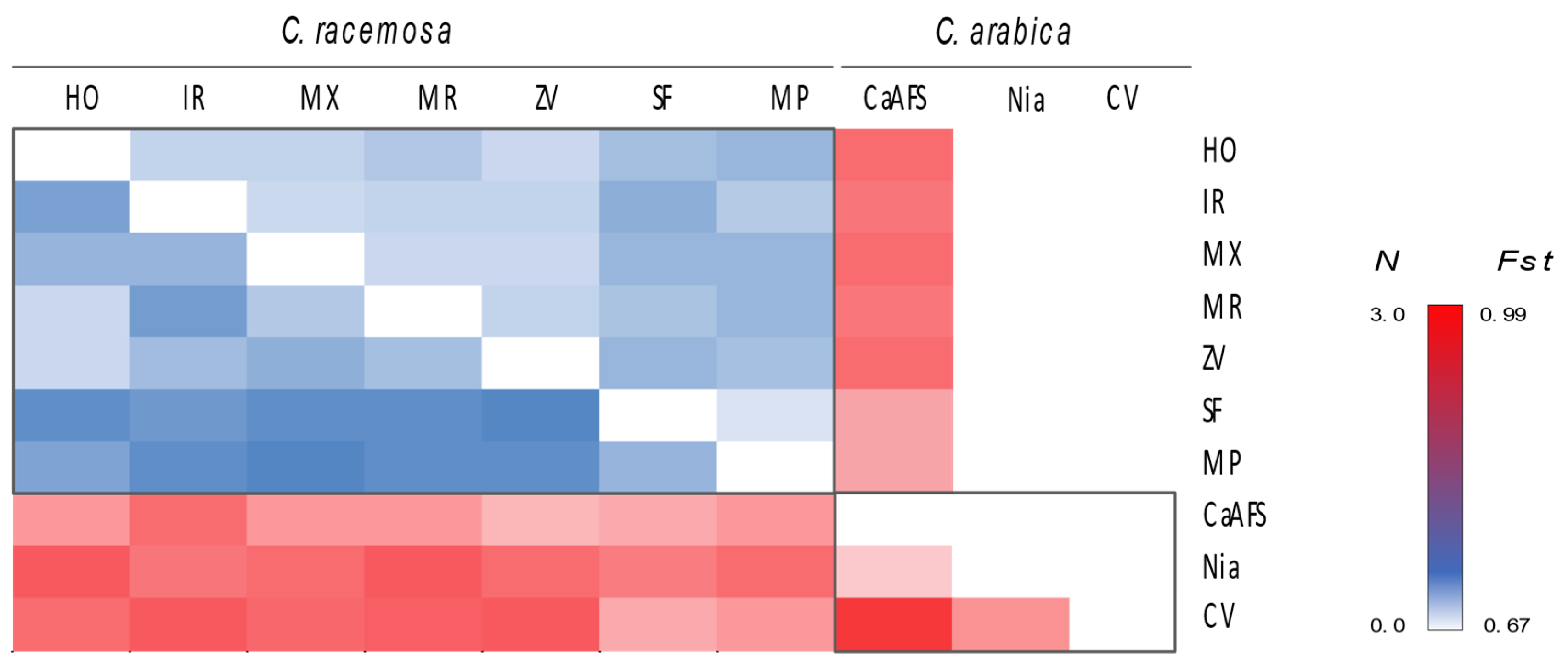
2.4. Genetic Differentiation between Species and Sites
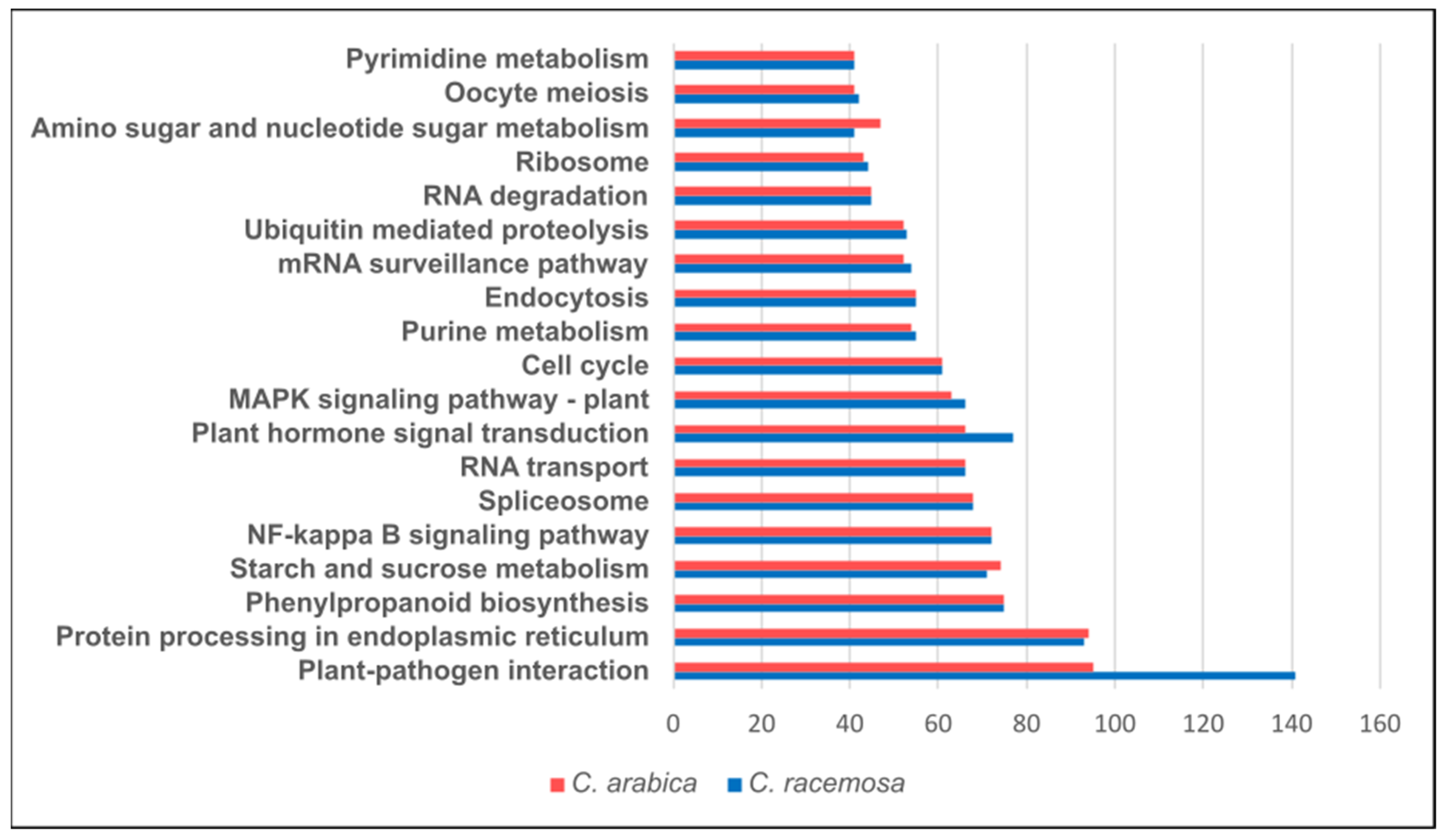
2.5. Annotation and Functional Pathways of SNPs
3. Discussion
3.1. Assessment of Coffea Species in Southern and Central Mozambique Using Plastid Markers
3.2. Low Genetic Diversity in Coffea arabica in Comparison with the Wild Relative C. racemosa
3.3. Implications for the Management of the Gorongosa Agroforestry System
4. Materials and Methods
4.1. Plant Sampling and DNA Isolation
4.2. Plastid Barcode Sequencing
4.3. Single-Sequence Polymorphic Repeats
4.4. GBS Library Preparation, Sequencing, and SNP Calling
4.5. Genetic Diversity, Structure, and Differentiation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ICO International Coffee Organization. What’s New. Available online: https://www.ico.org/ (accessed on 22 November 2022).
- DaMatta, F.M.; Cochicho Ramalho, J.D. Impacts of drought and temperature stress on coffee physiology and production: A review. Braz. J. Plant Physiol. 2006, 18, 55–81. [Google Scholar] [CrossRef]
- Davis, A.P.; Tosh, J.; Ruch, N.; Fay, M.F. Growing coffee: Psilanthus (Rubiaceae) subsumed on the basis of molecular and morphological data; implications for the size, morphology, distribution and evolutionary history of Coffea. Bot. J. Linn. Soc. 2011, 167, 357–377. [Google Scholar] [CrossRef]
- Dubberstein, D.; Lidon, F.C.; Rodrigues, A.P.; Semedo, J.N.; Marques, I.; Rodrigues, W.P.; Gouveia, D.; Armengaud, J.; Semedo, M.C.; Martins, S.; et al. Resilient and Sensitive Key Points of the Photosynthetic Machinery of Coffea spp. to the Single and Superimposed Exposure to Severe Drought and Heat Stresses. Front. Plant Sci. 2020, 11, 1049. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Marques, I.; Paulo, O.S.; Batista, D.; Partelli, F.L.; Lidon, F.C.; Damatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Understanding the impact of drought in Coffea genotypes: Transcriptomic analysis supports a common high resilience to moderate water deficit but a genotype dependent sensitivity to severe water deficit. Agronomy 2021, 11, 2255. [Google Scholar] [CrossRef]
- Venancio, L.P.; Filgueiras, R.; Mantovani, E.C.; do Amaral, C.H.; da Cunha, F.F.; dos Santos Silva, F.C.; Althoff, D.; dos Santos, R.A.; Cavatte, P.C. Impact of drought associated with high temperatures on Coffea canephora plantations: A case study in Espírito Santo State, Brazil. Sci. Rep. 2020, 10, 19719. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M.; Rahn, E.; Läderach, P.; Ghini, R.; Ramalho, J.C. Why could the coffee crop endure climate change and global warming to a greater extent than previously estimated? Clim. Chang. 2019, 152, 167–178. [Google Scholar] [CrossRef]
- de Oliveira, R.R.; Ribeiro, T.H.C.; Cardon, C.H.; Fedenia, L.; Maia, V.A.; Barbosa, B.C.F.; Caldeira, C.F.; Klein, P.E.; Chalfun-Junior, A. Elevated Temperatures Impose Transcriptional Constraints and Elicit Intraspecific Differences Between Coffee Genotypes. Front. Plant Sci. 2020, 11, 1113. [Google Scholar] [CrossRef] [PubMed]
- Byrareddy, V.; Kouadio, L.; Mushtaq, S.; Kath, J.; Stone, R. Coping with drought: Lessons learned from robusta coffee growers in Vietnam. Clim. Serv. 2021, 22, 100229. [Google Scholar] [CrossRef]
- Cassamo, C.T.; Draper, D.; Romeiras, M.M.; Marques, I.; Chiulele, R.; Rodrigues, M.; Stalmans, M.; Partelli, F.L.; Ribeiro-Barros, A.; Ramalho, J.C. Impact of climate changes in the suitable areas for Coffea arabica L. production in Mozambique: Agroforestry as an alternative management system to strengthen crop sustainability. Agric. Ecosyst. Environ. 2023, 346, 108341. [Google Scholar] [CrossRef]
- Läderach, P.; Ramirez–Villegas, J.; Navarro-Racines, C.; Zelaya, C.; Martinez–Valle, A.; Jarvis, A. Climate change adaptation of coffee production in space and time. Clim. Chang. 2017, 141, 47–62. [Google Scholar] [CrossRef]
- Chemura, A.; Mudereri, B.T.; Yalew, A.W.; Gornott, C. Climate change and specialty coffee potential in Ethiopia. Sci. Rep. 2021, 11, 8097. [Google Scholar] [CrossRef]
- Lara-Estrada, L.; Rasche, L.; Schneider, U.A. Land in Central America will become less suitable for coffee cultivation under climate change. Reg. Environ. Chang. 2021, 21, 88. [Google Scholar] [CrossRef]
- Scalabrin, S.; Toniutti, L.; Di Gaspero, G.; Scaglione, D.; Magris, G.; Vidotto, M.; Pinosio, S.; Cattonaro, F.; Magni, F.; Jurman, I.; et al. A single polyploidization event at the origin of the tetraploid genome of Coffea arabica is responsible for the extremely low genetic variation in wild and cultivated germplasm. Sci. Rep. 2020, 10, 4642. [Google Scholar] [CrossRef]
- Silvestrini, M.; Junqueira, M.G.; Favarin, A.C.; Guerreiro-Filho, O.; Maluf, M.P.; Silvarolla, M.B.; Colombo, C.A. Genetic diversity and structure of Ethiopian, Yemen and Brazilian Coffea arabica L. accessions using microsatellites markers. Genet. Resour. Crop Evol. 2007, 54, 1367–1379. [Google Scholar] [CrossRef]
- Jaramillo, J.; Muchugu, E.; Vega, F.E.; Davis, A.; Borgemeister, C.; Chabi-Olaye, A. Some like it hot: The influence and implications of climate change on coffee berry borer (Hypothenemus hampei) and coffee production in East Africa. PLoS ONE 2011, 6, e24528. [Google Scholar] [CrossRef]
- Davis, A.P.; Chester, M.; Maurin, O.; Fay, M.F. Searching for the relatives of Coffea (Rubiaceae, Ixoroideae): The circumscription and phylogeny of Coffeeae based on plastid sequence data and morphology. Am. J. Bot. 2007, 94, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Chadburn, H.; Moat, J.; O’Sullivan, R.; Hargreaves, S.; Lughadha, E.N. High extinction risk for wild coffee species and implications for coffee sector sustainability. Sci. Adv. 2019, 5, 3473–3489. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Gargiulo, R.; Almeida, I.N.d.M.; Caravela, M.I.; Denison, C.; Moat, J. Hot Coffee: The Identity, Climate Profiles, Agronomy, and Beverage Characteristics of Coffea racemosa and C. zanguebariae. Front. Sustain. Food Syst. 2021, 5, 383. [Google Scholar] [CrossRef]
- Halle, F.; Faria, M.T. Le Coffea racemosa Lour. Agron. Moçamb 1973, 7, 243–250. [Google Scholar]
- Guerreiro Filho, O. Coffea racemosa Lour. A Review. Café Cacao 1992, 36, 171–186. [Google Scholar]
- Davis, A.P.; Mieulet, D.; Moat, J.; Sarmu, D.; Haggar, J. Arabica-like flavour in a heat-tolerant wild coffee species. Nat. Plants 2021, 7, 413–418. [Google Scholar] [CrossRef]
- Davis, A.P.; Gole, T.W.; Baena, S.; Moat, J. The Impact of Climate Change on Indigenous Arabica Coffee (Coffea arabica): Predicting Future Trends and Identifying Priorities. PLoS ONE 2012, 7, e47981. [Google Scholar] [CrossRef]
- Gomes, L.C.; Bianchi, F.J.J.A.; Cardoso, I.M.; Fernandes, R.B.A.; Filho, E.I.F.; Schulte, R.P.O. Agroforestry systems can mitigate the impacts of climate change on coffee production: A spatially explicit assessment in Brazil. Agric. Ecosyst. Environ. 2020, 294, 106858. [Google Scholar] [CrossRef]
- Gidey, T.; Oliveira, T.S.; Crous-Duran, J.; Palma, J.H.N. Using the yield-SAFE model to assess the impacts of climate change on yield of coffee (Coffea arabica L.) under agroforestry and monoculture systems. Agrofor. Syst. 2020, 94, 57–70. [Google Scholar] [CrossRef]
- Cassamo, C.T.; Mangueze, A.V.J.; Leitão, A.E.; Pais, I.P.; Moreira, R.; Campa, C.; Chiulele, R.; Reis, F.O.; Marques, I.; Scotti-Campos, P.; et al. Shade and Altitude Implications on the Physical and Chemical Attributes of Green Coffee Beans from Gorongosa Mountain, Mozambique. Agronomy 2022, 12, 2540. [Google Scholar] [CrossRef]
- Koutouleas, A.; Sarzynski, T.; Bordeaux, M.; Bosselmann, A.S.; Campa, C.; Etienne, H.; Turreira-García, N.; Rigal, C.; Vaast, P.; Ramalho, J.C.; et al. Shaded-Coffee: A Nature-Based Strategy for Coffee Production Under Climate Change? A Review. Front. Sustain. Food Syst. 2022, 6, 158. [Google Scholar] [CrossRef]
- Koutouleas, A.; Sarzynski, T.; Bertrand, B.; Bordeaux, M.; Bosselmann, A.S.; Campa, C.; Etienne, H.; Turreira-García, N.; Léran, S.; Markussen, B.; et al. Shade effects on yield across different Coffea arabica cultivars—How much is too much? A meta-analysis. Agron. Sustain. Dev. 2022, 42, 55. [Google Scholar] [CrossRef]
- De Leijster, V.; Santos, M.J.; Wassen, M.W.; Camargo García, J.C.; Llorca Fernandez, I.; Verkuil, L.; Scheper, A.; Steenhuis, M.; Verweij, P.A. Ecosystem services trajectories in coffee agroforestry in Colombia over 40 years. Ecosyst. Serv. 2021, 48, 101246. [Google Scholar] [CrossRef]
- Nesper, M.; Kueffer, C.; Krishnan, S.; Kushalappa, C.G.; Ghazoul, J. Shade tree diversity enhances coffee production and quality in agroforestry systems in the Western Ghats. Agric. Ecosyst. Environ. 2017, 247, 172–181. [Google Scholar] [CrossRef]
- Sitoe, A.; Salomão, A.; Wertz-Kanounnikff, S. The Context of REDD+ in Mozambique Drivers, Agents and Institutions; CIFOR: Nairobi, Kenya, 2012. [Google Scholar]
- Hiern, W.P. On the African Species of the Genus Coffea, Linn. Trans. Linn. Soc. Lond. 2nd Ser. Bot. 1876, 1, 169–176. [Google Scholar] [CrossRef]
- Bridson, D.M. Coffea. In Flora of Tropical East Africa, Rubiaceae; Polhill, R.M., Bridson, D.M., Verdcourt, B., Eds.; Brookfield; Balkema: Rotterdam, The Netherlands, 1998; pp. 703–723. [Google Scholar]
- Davis, A.P.; Govaerts, R.; Bridson, D.M.; Ruhsam, M.; Moat, J.; Brummitt, N.A. A Global assessment of distribution, diversity, endemism, and taxonomic effort in the rubiaceae. Ann. Mo. Bot. Gard. 2009, 96, 68–78. [Google Scholar] [CrossRef]
- Bridson, D.M. Coffea. In Flora Zambesiaca; Pope, G.V., Ed.; Royal Botanic Gardens Kew: London, UK, 2003; pp. 452–463. [Google Scholar]
- Davis, A.P.; Govaerts, R.; Bridson, D.M.; Stoffelen, P. An annotated taxonomic conspectus of the genus Coffea (Rubiaceae). Bot. J. Linn. Soc. 2006, 152, 465–512. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- INFORM INFORM—Global, Open-Source Risk Assessment for Humanitarian Crises and Disasters. Available online: https://drmkc.jrc.ec.europa.eu/inform-index (accessed on 10 March 2023).
- Os Desastres Naturais em Moçambique Desde as Cheias de 2000—SIC Notícias. Available online: https://sicnoticias.pt/arquivo/ciclone-idai/2019-03-22-Os-desastres-naturais-em-Mocambique-desde-as-cheias-de-2000 (accessed on 10 March 2023).
- Mozambique|ReliefWeb. Available online: https://reliefweb.int/country/moz (accessed on 10 March 2023).
- Clovis, G.; Telina, R.; Frederique, M. Land Use and Land Cover Change 2000–2016 in Mozambique. Zenodo 2020. [Google Scholar] [CrossRef]
- Anthony, F.; Combes, M.C.; Astorga, C.; Bertrand, B.; Graziosi, G.; Lashermes, P. The origin of cultivated Coffea arabica L. varieties revealed by AFLP and SSR markers. Theor. Appl. Genet. 2002, 104, 894–900. [Google Scholar] [CrossRef]
- Maluf, M.P.; Silvestrini, M.; de Campos Ruggiero, L.M.; Guerreiro Filho, O.; Colombo, C.A. Genetic diversity of cultivated Coffea arabica inbred lines assessed by RAPD, AFLP and SSR marker systems. Sci. Agric. 2005, 62, 366–373. [Google Scholar] [CrossRef]
- Moncada, P.; McCouch, S. Simple sequence repeat diversity in diploid and tetraploid Coffea species. Genome 2004, 47, 501–509. [Google Scholar] [CrossRef]
- Dida, G.; Bantte, K.; Disasa, T. Molecular characterization of Arabica Coffee (Coffea arabica L.) germplasms and their contribution to biodiversity in Ethiopia. Plant Biotechnol. Rep. 2021, 15, 791–804. [Google Scholar] [CrossRef]
- Combes, M.C.; Andrzejewski, S.; Anthony, F.; Bertrand, B.; Rovelli, P.; Graziosi, G.; Lashermes, P. Characterization of microsatellite loci in Coffea arabica and related coffee species. Mol. Ecol. 2000, 9, 1178–1180. [Google Scholar] [CrossRef]
- Aggarwal, R.K.; Hendre, P.S.; Varshney, R.K.; Bhat, P.R.; Krishnakumar, V.; Singh, L. Identification, characterization and utilization of EST-derived genic microsatellite markers for genome analyses of coffee and related species. Theor. Appl. Genet. 2007, 114, 359–372. [Google Scholar] [CrossRef]
- Aerts, R.; Berecha, G.; Gijbels, P.; Hundera, K.; Van Glabeke, S.; Vandepitte, K.; Muys, B.; Roldán-Ruiz, I.; Honnay, O. Genetic variation and risks of introgression in the wild Coffea arabica gene pool in south-western Ethiopian montane rainforests. Evol. Appl. 2013, 6, 243–252. [Google Scholar] [CrossRef]
- Gadissa, F.; Tesfaye, K.; Dagne, K.; Geleta, M. Genetic diversity and population structure analyses of Plectranthus edulis (Vatke) Agnew collections from diverse agro-ecologies in Ethiopia using newly developed EST-SSRs marker system. BMC Genet. 2018, 19, 92. [Google Scholar] [CrossRef] [PubMed]
- Filho, H.P.M.; Carvalho, A.; Medina, D.M. Germoplasma de Coffea racemosa e seu potencial de melhoramento do cafeeiro. Bragantia 1977, 36, 43–46. [Google Scholar] [CrossRef]
- Senkoro, A.M.; Talhinhas, P.; Simões, F.; Batista-Santos, P.; Shackleton, C.M.; Voeks, R.A.; Marques, I.; Ribeiro-Barros, A.I. The genetic legacy of fragmentation and overexploitation in the threatened medicinal African pepper-bark tree, Warburgia salutaris. Sci. Rep. 2020, 10, 19725. [Google Scholar] [CrossRef]
- Marques, I.; Draper, D.; Riofrío, L.; Naranjo, C. Early Signs of the Effects of Forest Fragmentation on the Genetic Diversity and Structure of the Threatened Ecuadorian Tree Ocotea rotundata (Lauraceae). Forests 2022, 13, 1940. [Google Scholar] [CrossRef]
- Li, S.; Gan, X.; Han, H.; Zhang, X.; Tian, Z. Low within-population genetic diversity and high genetic differentiation among populations of the endangered plant Tetracentron sinense Oliver revealed by inter-simple sequence repeat analysis. Ann. For. Sci. 2018, 75, 74. [Google Scholar] [CrossRef]
- Husband, B.C. The role of triploid hybrids in the evolutionary dynamics of mixed-ploidy populations. Biol. J. Linn. Soc. 2004, 82, 537–546. [Google Scholar] [CrossRef]
- Botero, D.; Alvarado, C.; Bernal, A.; Danies, G.; Restrepo, S. Network analyses in plant pathogens. Front. Microbiol. 2018, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Jia, X.; Wu, Y.; Hu, Y.; Cheng, L.; Zhao, T.; Huang, Z.; Wang, Y. Quantitative proteomic analysis of Malus halliana exposed to salt-alkali mixed stress reveals alterations in energy metabolism and stress regulation. Plant Growth Regul. 2020, 90, 205–222. [Google Scholar] [CrossRef]
- Hamon, P.; Grover, C.E.; Davis, A.P.; Rakotomalala, J.-J.; Raharimalala, N.E.; Albert, V.A.; Sreenath, H.L.; Stoffelen, P.; Mitchell, S.E.; Couturon, E.; et al. Genotyping-by-sequencing provides the first well-resolved phylogeny for coffee (Coffea) and insights into the evolution of caffeine content in its species: GBS coffee phylogeny and the evolution of caffeine content. Mol. Phylogenet. Evol. 2017, 109, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Laíns e Silva, H.J. Subsídios Para o Fomento da Cultura do Café em Moçambique. Agência Geral do Ultramar; Divisão de Publicações: Lisboa, Portugal, 1954. [Google Scholar]
- Jezeer, R.E.; Verweij, P.A.; Santos, M.J.; Boot, R.G. Shaded Coffee and Cocoa—Double Dividend for Biodiversity and Small-scale Farmers. Ecol. Econ. 2017, 140, 136–145. [Google Scholar] [CrossRef]
- Cubry, P.; Pascal, M.; Hyacinte, L.; David, P.; de Bellis, F.; Poncet, V.; Anthony, F.; Dufour, M.; Leroy, T. Diversity in coffee assessed with SSR markers: Structure of the genus Coffea and perspectives for breeding. Genome 2008, 51, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Silva, H.L. Número de cromossomos em Coffea racemosa Lour. Bragantia 1956, 15, XVII–XVIII. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: Http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 15 May 2023).
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Depristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; Del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Rosenberg, N.A.; Donnelly, P. Association mapping in structured populations. Am. J. Hum. Genet. 2000, 67, 170–181. [Google Scholar] [CrossRef]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Cockerham, C.C. Variance of Gene Frequencies. Evolution 1969, 23, 72–84. [Google Scholar] [CrossRef]
- Keenan, K.; Mcginnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. diveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2014, e281. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sites | Na | Ne | I | Ho | He | F |
|---|---|---|---|---|---|---|---|
| C. racemosa | HO | 3.00 ± 0.21 | 2.59 ± 0.17 | 0.98 ± 0.06 | 0.93 ± 0.03 | 0.59 ± 0.02 | −0.58 ± 0.07 |
| IR | 2.71 ± 0.27 | 2.33 ± 0.19 | 0.85 ± 0.09 | 0.89 ± 0.07 | 0.53 ± 0.05 | −0.71 ± 0.08 | |
| MX | 2.71 ± 0.19 | 2.33 ± 0.14 | 0.87 ± 0.07 | 0.89 ± 0.07 | 0.54 ± 0.04 | −0.63 ± 0.11 | |
| MR | 2.86 ± 0.18 | 2.34 ± 0.13 | 0.89 ± 0.07 | 0.86 ± 0.07 | 0.55 ± 0.03 | −0.55 ± 0.10 | |
| ZV | 3.00 ± 0.26 | 2.47 ± 0.18 | 0.94 ± 0.07 | 0.89 ± 0.05 | 0.57 ± 0.03 | −0.56 ± 0.08 | |
| SF | 2.93 ± 0.22 | 2.45 ± 0.20 | 0.91 ± 0.08 | 0.89 ± 0.06 | 0.55 ± 0.04 | −0.59 ± 0.08 | |
| MP | 2.92 ± 0.29 | 2.36 ± 0.15 | 0.90 ± 0.07 | 0.95 ± 0.03 | 0.56 ± 0.02 | −0.72 ± 0.08 | |
| average | 2.88 ± 0.09 | 2.41 ± 0.06 | 0.91 ± 0.03 | 0.90 ± 0.02 | 0.56 ± 0.01 | −0.62 ± 0.03 | |
| C. arabica | CaAFS | 1.93 ± 0.17 | 1.26 ± 0.10 | 0.27 ± 0.06 | 0.16 ± 0.07 | 0.16 ± 0.05 | 0.02 ± 0.11 |
| Nia | 1.88 ± 0.16 | 1.80 ± 0.13 | 0.56 ± 0.09 | 0.69 ± 0.12 | 0.39 ± 0.06 | −0.80 ± 0.11 | |
| CV | 1.79 ± 0.24 | 1.73 ± 0.23 | 0.45 ± 0.13 | 0.05 ± 0.03 | 0.29 ± 0.08 | 0.83 ± 0.08 | |
| average | 1.87 ± 0.08 | 1.68 ± 0.07 | 0.48 ± 0.04 | 0.46 ± 0.05 | 0.39 ± 0.03 | −0.40 ± 0.09 | |
| All samples | 2.46 ± 0.08 | 2.10 ± 0.054 | 0.73 ± 0.03 | 0.72 ± 0.03 | 0.46 ± 0.01 | 0.55 ± 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tapaça, I.d.P.E.; Mavuque, L.; Corti, R.; Pedrazzani, S.; Maquia, I.S.A.; Tongai, C.; Partelli, F.L.; Ramalho, J.C.; Marques, I.; Ribeiro-Barros, A.I. Genomic Evaluation of Coffea arabica and Its Wild Relative Coffea racemosa in Mozambique: Settling Resilience Keys for the Coffee Crop in the Context of Climate Change. Plants 2023, 12, 2044. https://doi.org/10.3390/plants12102044
Tapaça IdPE, Mavuque L, Corti R, Pedrazzani S, Maquia ISA, Tongai C, Partelli FL, Ramalho JC, Marques I, Ribeiro-Barros AI. Genomic Evaluation of Coffea arabica and Its Wild Relative Coffea racemosa in Mozambique: Settling Resilience Keys for the Coffee Crop in the Context of Climate Change. Plants. 2023; 12(10):2044. https://doi.org/10.3390/plants12102044
Chicago/Turabian StyleTapaça, Inocência da Piedade Ernesto, Lopes Mavuque, Riccardo Corti, Samuele Pedrazzani, Ivete S. A. Maquia, Castigo Tongai, Fábio Luiz Partelli, José C. Ramalho, Isabel Marques, and Ana I. Ribeiro-Barros. 2023. "Genomic Evaluation of Coffea arabica and Its Wild Relative Coffea racemosa in Mozambique: Settling Resilience Keys for the Coffee Crop in the Context of Climate Change" Plants 12, no. 10: 2044. https://doi.org/10.3390/plants12102044