
Repeatedly Northwards and Upwards: Southern African Grasslands Fuel the Colonization of the African Sky Islands in Helichrysum (Compositae)
 , , , , , , , , , ,
, , , , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
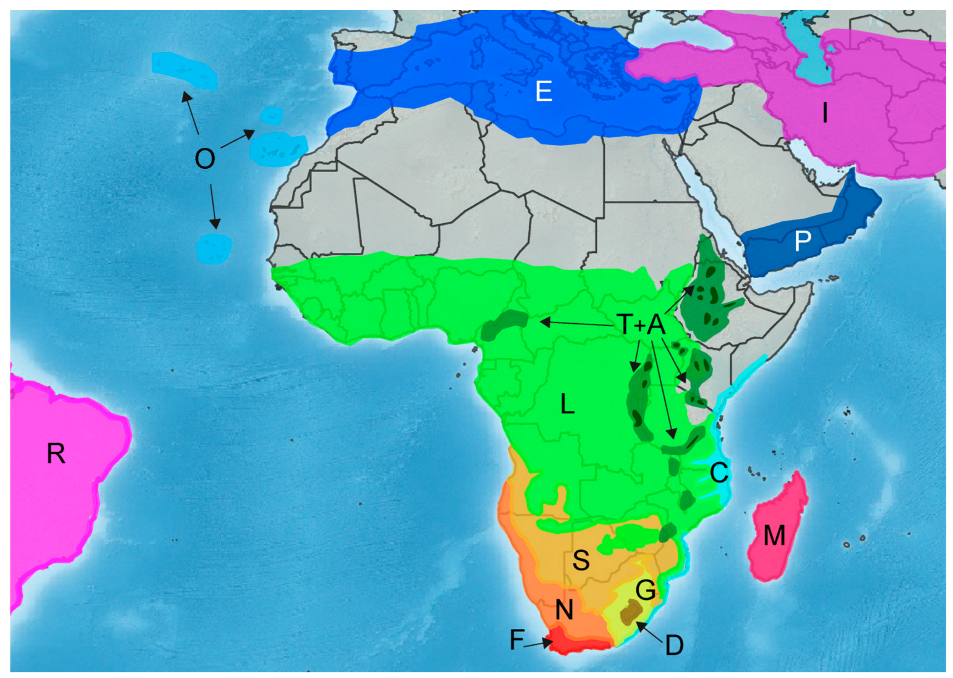
2.1. Taxon Sampling
2.2. DNA Extraction, Library Preparation, Target Capture, and Sequencing
2.3. Molecular Data Processing and Phylogenetic Analyses
2.4. Divergence Time Estimation
2.5. Ancestral Range Estimation
3. Results
3.1. Alignment Processing and Filtering
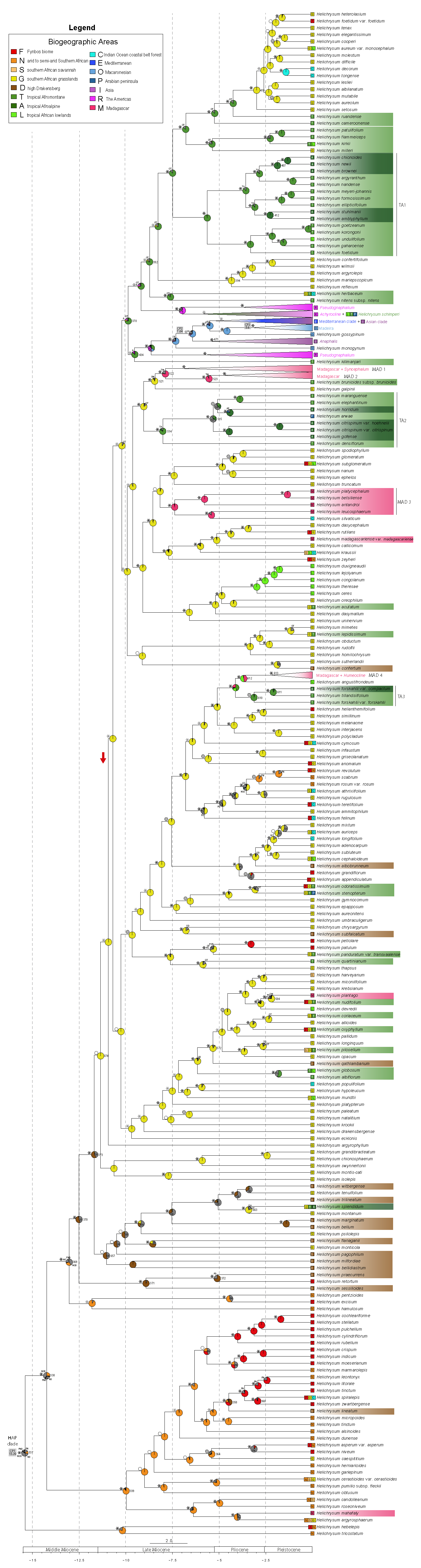
3.2. Phylogenetic Analyses
3.3. Divergence Time and Ancestral Range Estimation
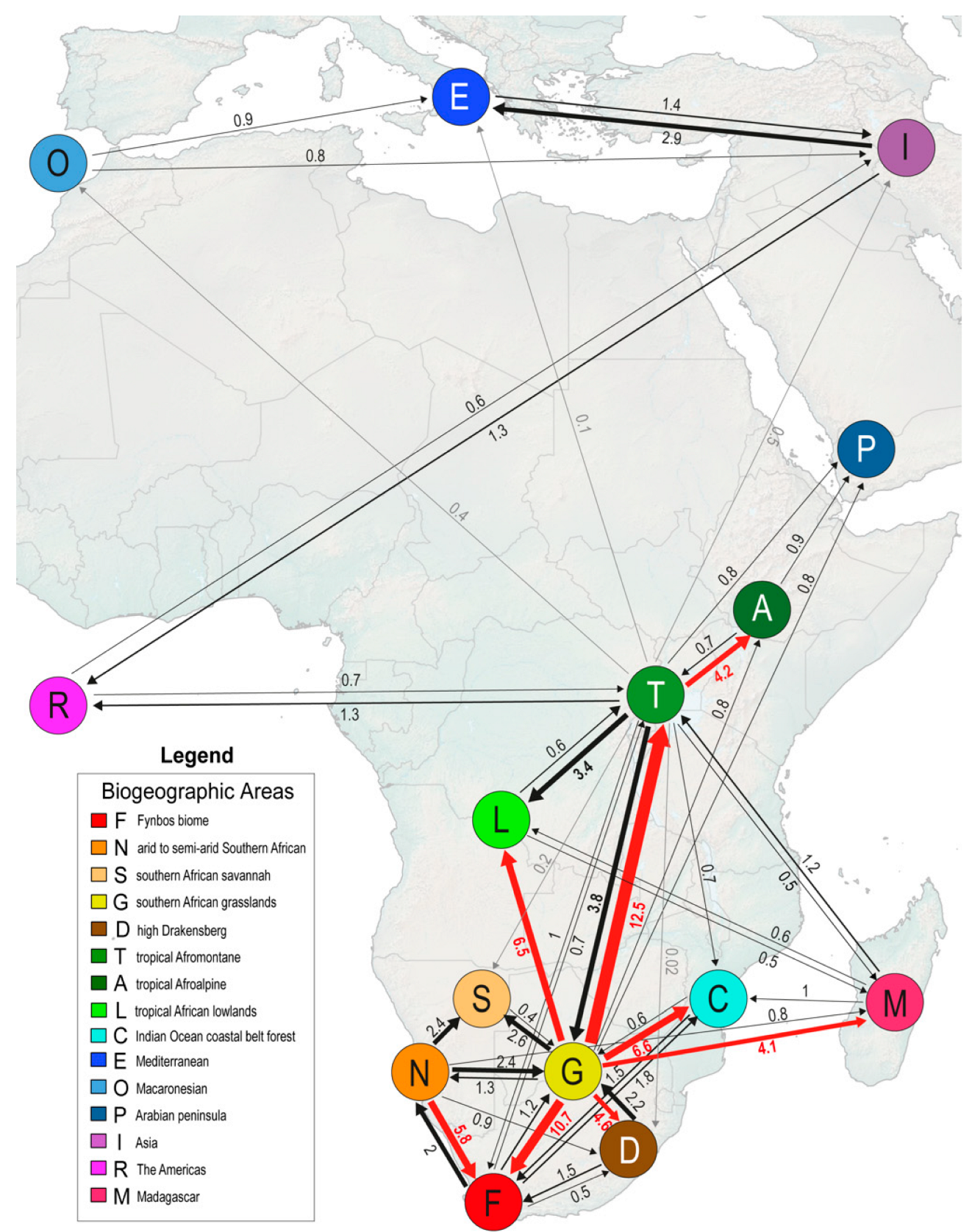
3.4. Number, Type, and Directionality Estimation of Biogeographical Events
4. Discussion
4.1. Utility of Target-Enrichment Strategies in Reconstructing the Radiation of Helichrysum
4.2. The Early History of Helichrysum and Colonization of Madagascar
4.3. Repeatedly Northwards
4.4. Repeatedly Upwards
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heald, W.F. Sky Island; Van Nostrand: Princeton, NJ, USA, 1967. [Google Scholar]
- White, F. The Afromontane region. In Biogeography and Ecology of Southern Africa; Wegner, M.J.A., Ed.; Springer: Dordrecht, The Netherlands, 1978; Volume 31, pp. 463–513. [Google Scholar] [CrossRef]
- White, F. Vegetation of Africa: A Descriptive Memoir to Accompany the UNESCO/AETFAT/UNSO Vegetation Map of Africa; UNESCO: Paris, France, 1983. [Google Scholar]
- Gehrke, B.; Linder, H.P. Species richness, endemism and species composition in the tropical Afroalpine flora. Alp. Bot. 2014, 124, 165–177. [Google Scholar] [CrossRef]
- Hedberg, O. Evolution and speciation in a tropical high mountain flora. Biol. J. Linn. Soc. 1969, 1, 135–148. [Google Scholar] [CrossRef]
- Rutherford, M.C.; Westfall, R.H. Biomes of southern Africa—An objective categorization. Memo. Bot. Surv. S. Afr. 1986, 54, 1–98. [Google Scholar]
- Davis, S.D.; Heywood, V.H.; Hamilton, A.C. Centres of plant diversity. In A Guide and Strategy for their Conservation. Europe, Africa, South West Asia and the Middle East; IUCN Publications Unit: Cambridge, UK, 1994; Volume 1. [Google Scholar]
- Burgoyne, P.M.; Van Wyk, A.E.; Anderson, J.M.; Schrire, B.D. Phanerozoic evolution of plants on the African Plate. J. Afr. Earth Sci. 2005, 43, 13–52. [Google Scholar] [CrossRef]
- Carbutt, C.; Edwards, T.J. Reconciling ecological and phytogeographical spatial boundaries to clarify the limits of the montane and alpine regions of sub-Sahelian Africa. S. Afr. J. Bot. 2015, 98, 64–75. [Google Scholar] [CrossRef]
- Gehrke, B.; Linder, H.P. The scramble for Africa Pan-temperate elements on the African high mountains. Proc. R. Soc. B: Biol. Sci. 2009, 276, 2657–2665. [Google Scholar] [CrossRef] [Green Version]
- Kedebe, M.; Ehrich, D.; Taberlet, P.; Nemomissa, S.; Brochmann, C. Phylogeography and conservation genetics of a giant lobelia (Lobelia giberroa) in Ethiopian and Tropical East Africa Mountains. Mol. Ecol. 2007, 16, 1233–1243. [Google Scholar] [CrossRef]
- Mairal, M.; Pokorny, L.; Aldasoro, J.J.; Alarcón, M.; Sanmartín, I. Ancient vicariance and climate-driven extinction explain continental-wide disjunctions in Africa: The case of the Rand Flora genus Canarina (Campanulaceae). Mol. Ecol. 2015, 24, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Hemp, A. Continuum or zonation? Altitudinal gradients in the forest vegetation of Mt. Kilimanjaro. Plant Ecol. 2006, 184, 27–42. [Google Scholar] [CrossRef]
- Kandziora, M.; Gehrke, B.; Popp, M.; Gizaw, A.; Brochmann, C.; Pirie, M.D. The enigmatic tropical alpine flora on the African sky islands is young, disturbed, and unsaturated. Proc. Natl. Acad. Sci. USA 2022, 119, e2112737119. [Google Scholar] [CrossRef]
- Engler, A. Plants of the northern temperate zone in their transition to the high mountains of tropical Africa. Ann. Bot. 1904, 18, 523–540. [Google Scholar] [CrossRef]
- Killick, D.J.B. The Afro-Alpine region. In Biogeography and Ecology of Southern Africa; Wegner, M.J.A., Ed.; Monographiae Biologicae: The Hague, The Netherlands, 1978; Volume 31, pp. 515–542. [Google Scholar] [CrossRef]
- White, F. Long-distance dispersal and the origins of the Afromontane flora. Sonderbd. Des Nat. Ver. Hambg. 1983, 7, 87–116. [Google Scholar]
- Galley, C.; Bytebier, B.; Bellstedt, D.U.; Linder, H.P. The Cape element in the Afrotemperate flora: From Cape to Cairo? Proc. R. Soc. B Biol. Sci. 2007, 274, 535–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, J.; Verboom, G.A.; Bergh, N.G. Erosive processes after tectonic uplift stimulate vicariant and adaptive speciation: Evolution in an Afrotemperate-endemic paper daisy genus. BMC Evol. Biol. 2014, 14, 27. [Google Scholar] [CrossRef] [Green Version]
- Brochmann, C.; Gizaw, A.; Chala, D.; Kandziora, M.; Eilu, G.; Popp, M.; Pirie, M.D.; Gehrke, B. History and evolution of the afroalpine flora: In the footsteps of Olov Hedberg. Alp. Bot. 2021, 132, 65–87. [Google Scholar] [CrossRef]
- Chala, D.; Zimmermann, N.E.; Brochmann, C.; Bakkesstuen, V. Migration corridors for alpine plants among the “sky islands” of eastern Africa: Do they, or did they exist? Alp. Bot. 2017, 127, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Popp, M.; Gizaw, A.; Nemomissa, S.; Suda, J.; Brochmann, C. Colonization and diversification in the African “sky islands” by Eurasian Lychnis L. (Caryophyllaceae). J. Biogeogr. 2008, 35, 1016–1029. [Google Scholar] [CrossRef]
- Kandziora, M.; Kadereit, J.W.; Gehrke, B. Frequent colonization and little in situ speciation in Senecio in the tropical alpine-like islands of eastern Africa. Am. J. Bot. 2016, 103, 1483–1498. [Google Scholar] [CrossRef] [Green Version]
- Galbany-Casals, M.; Unwin, M.; Garcia-Jacas, N.; Smissen, R.D.; Susanna, A.; Bayer, R.J. Phylogenetic relationships in Helichrysum (Compositae: Gnaphalieae) and related genera: Incongruence between nuclear and plastid phylogenies, biogeographic and morphological patterns, and implications for generic delimitation. Taxon 2014, 63, 608–624. [Google Scholar] [CrossRef] [Green Version]
- Gehrke, B. Staying cool: Preadaptation to temperate climates required for colonizing tropical alpine-like environments. PhytoKeys 2018, 96, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Hilliard, O.M. Helichrysum Mill. In Flora of Southern Africa; Leistner, O.A., Ed.; Department of Agriculture: Pretoria, South Africa, 1983; Volume 33, pp. 61–310, Part 7, Fascicle 2. [Google Scholar]
- Anderberg, A.A. Taxonomy and phylogeny of the tribe Gnaphalieae (Asteraceae). Opera Bot. 1991, 104, 1–195. [Google Scholar]
- Galbany-Casals, M.; Garcia-Jacas, N.; Sáez, L.; Benedí, C.; Susanna, A. Phylogeny, Biogeography and character evolution in Mediterranean, Asiatic and Macaronesian Helichrysum (Asteraceae, Gnaphalieae) inferred from nuclear phylogenetic analyses. Int. J. Plant. Sci. 2009, 170, 365–380. [Google Scholar] [CrossRef] [Green Version]
- Carbutt, C. The Drakensberg Mountain Centre: A necessary revision of southern Africa’s high-elevation centre of plant endemism. S. Afr. J. Bot. 2019, 124, 508–529. [Google Scholar] [CrossRef]
- Humbert, H. Flore de Madagascar et des Comores (Plantes vasculaires); 189e famille. Composées, part 2; Leroy, J.F., Ed.; Muséum National d’Histoire Naturelle: Paris, France, 1962. [Google Scholar]
- Andrés-Sánchez, S.; Verboom, G.A.; Galbany-Casals, M.; Bergh, N.G. Evolutionary history of the arid climate-adapted Helichrysum (Asteraceae: Gnaphalieae): Cape origin and association between annual life-history and low chromosome numbers. J. Syst. Evol. 2019, 57, 468–487. [Google Scholar] [CrossRef]
- Cronn, R.; Knaus, B.J.; Liston, A.; Maughan, P.J.; Parks, M.; Syring, J.V.; Undall, J. Targeted enrichment strategies for next-generation plant biology. Am. J. Bot. 2012, 99, 291–311. [Google Scholar] [CrossRef] [Green Version]
- Mandel, J.R.; Dikow, R.B.; Funk, V.A.; Masalia, R.R.; Staton, S.E.; Kozik, A.; Michelmore, R.W.; Rieseberg, L.H.; Burke, J.M. A target enrichment method for gathering phylogenetic information from hundreds of loci: An example from the Compositae. Appl. Plant Sci. 2014, 2, 1300085. [Google Scholar] [CrossRef]
- Ufimov, R.; Gorospe, J.M.; Fér, T.; Kandziora, M.; Salomon, L.; van Loo, M.; Schmickl, R. Utilizing paralogues for phylogenetic reconstruction has the potential to increase species tree support and reduce gene tree discordance in target enrichment data. Mol. Ecol. Resour. 2022, 22, 3018–3034. [Google Scholar] [CrossRef]
- Smissen, R.D.; Bayer, R.J.; Berg, N.G.; Breitwieser, I.; Freire, S.E.; Galbany-Casals, M.; Schmidt-Lebuhn, A.N.; Ward, J.M. A revised subtribal classification of Gnaphalieae (Asteraceae). Taxon 2020, 6, 778–806. [Google Scholar] [CrossRef]
- Nie, Z.-L.; Funk, V.A.; Meng, Y.; Deng, T.; Sun, H.; Wen, J. Recent assembly of the global herbaceous flora: Evidence from the paper daisies (Asteraceae: Gnaphalieae). New Phytol. 2015, 209, 1789–1806. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. The New York Botanical Garden, Bronx. Available online: http://sweetgum.nybg.org/ih/ (accessed on 20 April 2021).
- Adams, C.D. Compositae. In Flora of West Tropical Africa, 2nd ed.; Hutchinson, J., Dalziel, J.M., Keay, R.W.Y., Hepper, F.N., Eds.; with annotations from African Plants Database (Version 3.4.0); Crown Agents: London, UK, 1963; Volume 2, pp. 225–297. [Google Scholar]
- Beentje, H.J. Helichrysum. In Flora of tropical East Africa; part 2; Beentje, H.J., Ed.; Balkema: Rotterdam, The Netherlands, 2002; pp. 403–452. [Google Scholar]
- Bingham, M.G.; Willemen, A.; Wursten, B.T.; Ballings, P.; Hyde, M.A. Flora of Zambia, Based on Flora Zambesiaca. Available online: http://www.zambiaflora.com (accessed on 21 April 2021).
- Carbutt, C.; Edwards, T.J. The flora of the Drakensberg Alpine Centre. Edinburgh J. Bot. 2004, 60, 581–607. [Google Scholar] [CrossRef]
- Guillarmod, A.J. Flora of Lesotho (Basutoland); Verlag von Cramer: Lehre, Germany, 1971. [Google Scholar]
- Friis, I.; Vollesen, K. Flora of the Sudan-Uganda border area east of the Nile. In Catalogue of Vascular Plants; 1st part; Kongel Danske Vidensk Selsk: Copenhagen, Denmark, 1998; Volume 1. [Google Scholar] [CrossRef]
- Hyde, M.A.; Wursten, B.T.; Ballings, P.; Coates Palgrave, M. Flora of Botswana, Based on Flora Zambesiaca. Kew Royal Botanic Gardens. Available online: http://www.botswanaflora.com/index.php (accessed on 22 April 2021).
- Hyde, M.A.; Wursten, B.T.; Ballings, P.; Coates Palgrave, M. Flora of Malawi, Based on Flora Zambesiaca. Kew Royal Botanic Gardens. Available online: http://www.malawiflora.com (accessed on 22 April 2021).
- Hyde, M.A.; Wursten, B.T.; Ballings, P.; Coates Palgrave, M. Flora of Mozambique, Based on Flora Zambesiaca. Kew Royal Botanic Gardens. Available online: http://www.mozambiqueflora.com (accessed on 23 April 2021).
- Hyde, M.A.; Wursten, B.T.; Ballings, P.; Coates Palgrave, M. Flora of Zimbabwe, Based on Flora Zambesiaca. Kew Royal Botanic Gardens. Available online: http://www.zimbabweflora.co.zw (accessed on 23 April 2021).
- Koekemoer, M.; Steyn, H.M. Helichrysums of Southern Africa, Version 1. South African National Biodiversity Institute. 2010. Available online: https://keys.lucidcentral.org/keys/v3/helichrysum/key/Helichrysum/Media/Html/home.htm (accessed on 15 April 2021).
- Lisowski, S. Helichrysum Compositae (deuxième partie: Tribu Inuleae). In Flore d’Afrique Centrale (Zaire, Rwanda, Burundi); National Botanic Garden of Belgium: Meise, Belgium, 1989; pp. 68–192. [Google Scholar]
- Tadesse, M. Helichrysum Mill., Asteraceae (Compositae). In Flora of Ethiopia and Eritrea; part 2; Addis Abeba University: Addis Ababa, Ethiopia, 2004; pp. 163–178. [Google Scholar]
- Threatened Species Programme. Red List of South African Plants. South African National Biodiversity Institute (2010–2012). Available online: http://redlist.sanbi.org/index.php (accessed on 20 April 2021).
- Wood, J.R.I. A Handbook of the Yemen Flora; Kew Publishing: Kew, UK, 1997. [Google Scholar]
- Baker, M.S.; Kane, N.C.; Matvienko, M.; Kozik, A.; Michelmore, R.W.; Knapp, S.J.; Rieseberg, L.H. Multiple paleopolyploidizations during the evolution of the Compositae reveal parallel patterns of duplicate gene retention after millions of years. Mol. Biol. Evol. 2008, 25, 2445–2455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, M.S.; Li, Z.; Kidder, T.I.; Reardon, C.R.; Lai, Z.; Oliveira, L.O.; Scascitelli, M.; Rieserberg, L.H. Most Compositae (Asteraceae) are descendant of a paleohexaploid and all share a paleotetraploid ancestor with the Calyceraceae. Am. J. Bot. 2016, 103, 1203–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smissen, R.D.; Galbany-Casals, M.; Breitwieser, I. Ancient allopolyploidy in the everlasting daisies (Asteraceae: Gnaphalieae): Complex relationship among extant clades. Taxon 2011, 60, 649–662. [Google Scholar] [CrossRef]
- Fér, T.; Schmickl, R.E. HybPhyloMaker: Target Enrichment data analysis from raw reads to species trees. Evol. Bioinform. Online 2018, 14, 1176934317742613. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-AWARE Aligner; Lawrence Berkeley National Lab (LBNL): Berkeley, CA, USA, 2014. [Google Scholar]
- Kent, W.J. BLAT—the BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Fernández, J. A practical guide to design and assess a phylogenomic study. Genome Biol. Evol. 2022, 14, evac129. [Google Scholar] [CrossRef]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A new and scalable tool for the selection of DNA and protein evolutionary models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.; Domelevo Entfellner, J.B.; Wilkinson, E.; Correia, D.; Dávila Felipe, M.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Rabiee, M.; Sayyari, E.; Mirarab, S. ASTRAL-III: Polytomial time species tree reconstruction from partially resolved gene trees. BMC Bioinform. 2018, 19, 153. [Google Scholar] [CrossRef] [Green Version]
- Sayyari, E.; Mirarab, S. Fast coalescent-based computation of local branch support from quartet frequencies. Mol. Biol. Evol. 2016, 33, 1654–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree ver. 1.4.3. Department of Zoology, University of Oxford, Oxford. 2016. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 30 November 2021).
- Sanderson, M.J. Estimating absolute rates of molecular evolution and divergence times: A Penalized Likelihood approach. Mol. Biol. Evol. 2002, 19, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.A.; O’Meara, B.C. treePL: Divergence time estimation using penalized likelihood for large phylogenies. Bioinformatics 2012, 28, 2689–2690. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.Y.M.; Phillips, M.J. Accounting for Calibration Uncertainty in Phylogenetic Estimation of Evolutionary Divergence Times. Syst. Biol. 2009, 58, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Heath, T.A. Dating Species Divergences with the Fossilized Birth-Death Process. Divergence Time Estimation Using BEAST v2.x. 2020. Available online: http://phyloworks.org/workshops/DivTime_BEAST2_tutorial_FBD.pdf (accessed on 8 May 2023).
- Ramalho, R.S.; Brum da Silveira, A.; Fonseca, P.E.; Madeira, J.; Cosca, M.; Cachao, M.; Fonseca, M.M.; Prada, S.N. The emergence of volcanic oceanic islands on a slow-moving plate: The example of Madeira Island, NE Atlantic. Geochem. Geophys. Geosystems 2015, 16, 522–537. [Google Scholar] [CrossRef]
- Van Wyk, A.E.; Smith, G.F. Regions of Floristic Endemism in Southern Africa: A Review with Emphasis on Succulents; Umdaus Press: Hatfield, South Africa, 2001. [Google Scholar]
- Rutherford, M.C.; Mucina, L.; Powrie, L.W. Biomes and bioregions of Southern Africa. In The vegetation of South Africa, Lesotho and Swaziland; Mucina, L., Rutherford, M.C., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2006; pp. 30–51. [Google Scholar]
- Matzke, N.J. BioGeoBEARS: Biogeography with Bayesian (and Likelihood) Evolutionary Analysis in R Scripts. R package. 2013. Available online: https://github.com/nmatzke/BioGeoBEARS (accessed on 2 September 2021).
- Ree, R.H.; Moore, B.R.; Webb, C.O.; Donoghue, M.J. A likelihood framework for inferring the evolution of geographic range on phylogenetic trees. Evolution 2005, 59, 2299–2311. [Google Scholar] [CrossRef] [PubMed]
- Ree, R.H.; Smith, S.A. Maximum likelihood inference of geographic range evolution by dispersal, local extinction and cladogenesis. Syst. Biol. 2008, 57, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F. Dispersal–vicariance analysis: A new approach to the quantification of historical biogeography. Syst. Biol. 1997, 46, 195–203. [Google Scholar] [CrossRef]
- Landis, M.J.; Matzke, N.J.; Moore, B.R.; Huelsenbeck, J.P. Bayesian analysis of biogeography when the number of areas is large. Syst. Biol. 2013, 62, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Matzke, N.J. Model selection in historical biogeography reveals that founder-event speciation is a crucial process in island clades. Syst. Biol. 2014, 63, 951–970. [Google Scholar] [CrossRef] [PubMed]
- Dupin, J.; Matzke, N.J.; Särkinen, T.; Knapp, S.J.; Olmstead, R.G.; Bohs, L.; Smith, S.D. Bayesian estimation of the global biogeographical history of the Solanaceae. J. Biogeogr. 2017, 44, 887–899. [Google Scholar] [CrossRef]
- Ree, R.H.; Sanmartín, I. Conceptual and statistical problems with the DEC+J model of founder-event speciation and its comparison with DEC via model selection. J. Biogeogr. 2018, 45, 741–749. [Google Scholar] [CrossRef]
- Matzke, N.J. Statistical comparison of DEC and DEC+J is identical to comparison of two ClaSSE submodels, and is therefore valid. J. Biogeogr. 2022, 49, 1805–1824. [Google Scholar] [CrossRef]
- Seton, M.; Müller, R.D.; Zahirovic, S.; Gaina, C.; Torsvik, T.; Shephard, G.; Talsma, A.; Gurnis, M.; Turner, M.; Maus, S.; et al. Global continental and ocean basin reconstructions since 200 Ma. Earth Sci. Rev. 2012, 113, 212–270. [Google Scholar] [CrossRef] [Green Version]
- Yoder, A.D.; Nowak, M.D. Has vicariance or dispersal been the predominant biogeographic force in Madagascar? Only time will tell. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 404–431. [Google Scholar] [CrossRef] [Green Version]
- Weitemier, K.; Straub, S.C.; Cronn, R.C.; Fischbein, M.; Schmickl, R.; McDonnell, A.; Liston, A. Hyb-Seq: Combining target enrichment and genome skimming for plant phylogenomics. App. Plant Sci. 2014, 2, apps.1400042. [Google Scholar] [CrossRef] [PubMed]
- Bagley, J.C.; Uribe-Convers, S.; Carlsen, M.M.; Muchhala, N. Utility of targeted sequence capture for phylogenomics in rapid, recent angiosperm radiations: Neotropical Burmeistera bellflowers as a case study. Mol. Phylogenet. Evol. 2020, 152, 106769. [Google Scholar] [CrossRef] [PubMed]
- Gizaw, A.; Gorospe, J.M.; Kandziora, M.; Chala, D.; Gustafsoon, L.; Zinaw, A.; Salomón, L.; Eilu, G.; Brochmann, C.; Kolář, F.; et al. Afro-alpine flagships revisited II: Elucidating the evolutionary relationships and species boundaries in the giant senecios (Dendrosenecio, Asteraceae). Alp. Bot. 2021, 132, 89–105. [Google Scholar] [CrossRef]
- Lagomarsino, L.P.; Frankel, L.; Uribe-Convers, S.; Antonelli, A.; Muchhala, N. Increased resolution in the face of conflict: Phylogenomics of the Neotropical bellflowers (Campanulaceae: Lobelioideae), a rapid plant radiation. Ann. Bot. 2022, 129, 723–736. [Google Scholar] [CrossRef]
- Kandziora, M.; Sklenář, P.; Kolář, F.; Schmickl, R. How to tackle phylogenetic discordance in recent and rapidly radiating groups? Developing a workflow using Loricaria (Asteraceae) as an example. Front. Plant Sci. 2022, 12, 765719. [Google Scholar] [CrossRef]
- Whiterfield, J.B.; Lockhart, P.J. Deciphering ancient rapid radiations. Trends Ecol. Evol. 2007, 22, 258–265. [Google Scholar] [CrossRef]
- Sepulchre, P.; Ramstein, G.; Fluteau, F.; Schuster, M.; Tercelin, J.J.; Brunet, M. Tectonic uplift and Eastern Africa aridification. Science 2006, 313, 1419–1423. [Google Scholar] [CrossRef] [Green Version]
- Maddison, W.P. Gene trees in species trees. Syst. Biol. 1997, 46, 523–536. [Google Scholar] [CrossRef]
- Linkem, C.W.; Minin, V.N.; Leaché, A.D. Detecting the anomaly zone in species trees and evidence for a misleading signal in higher-level skink phylogeny (Squamata: Scincidae). Syst. Biol. 2016, 65, 465–477. [Google Scholar] [CrossRef] [Green Version]
- Westerhold, T.; Marwan, N.; Drury, A.J.; Liebrand, D.; Agnini, C.; Anagnostou, E.; Barnet, J.; Bohaty, S.M.; De Vleeschouwer, D.; Florindo, F.; et al. An astronomically dated record of Earth’s climate and its predictability over the last 66 million years. Science 2020, 369, 1383–1387. [Google Scholar] [CrossRef]
- Coetzee, J.A.; Rogers, J. Palynological and lithological evidence for the Miocene palaeoenvironment in the Saldanha region (South Africa). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1982, 39, 71–85. [Google Scholar] [CrossRef]
- Dupont, L.M.; Linder, H.P.; Rommerskirchen, F.; Schefuß, E. Climate driven rampant speciation of the Cape flora. J. Biogeogr. 2011, 38, 1059–1068. [Google Scholar] [CrossRef]
- Partridge, T.C.; Maud, R.R. Geomorphic evolution of southern Africa since the Mesozoic. S. Afr. Geogr. J. 1987, 90, 179–208. [Google Scholar] [CrossRef]
- Partridge, T.C. Of diamonds, dinosaurs and diastrophism: 150 years of landscape evolution in southern Africa. S. Afr. Geogr. J. 1998, 101, 167–184. [Google Scholar] [CrossRef]
- Partridge, T.C.; Maud, R.R. Macro-scale geomorphic evolution of southern Africa. Oxf. Monogr. Geol. Geophys. 2000, 40, 3–18. [Google Scholar]
- Hughes, C.E.; Atchison, G.W. The Ubiquity of alpine plant radiations: From the Andes to the Hengduan Mountains. New Phytol. 2015, 207, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Mikula, O.; Nicolas, V.; Šumbera, R.; Konečný, A.; Deneys, C.; Verheyen, E.; Bryjová, A.; Lemmon, A.R.; Lemmon, E.M.; Bryja, J. Nuclear phylogenomics, but not mitogenomics, resolves the most successful Late Miocene radiation of African mammals (Rodentia: Muridae: Arvicanthini). Mol. Phylogenet. Evol. 2021, 157, 107069. [Google Scholar] [CrossRef]
- Aduse-Poku, K.; van Bergen, E.; Sáfián, S.; Collins, S.C.; Etienne, R.S.; Herrera-Alsina, L.; Brakefield, P.M.; Brattström, O.; Lohman, D.J.; Wahlberg, N. Miocene climate and habitat change drove diversification in Bicyclus, Africa’s largest radiation of Satyrine butterflies. Syst. Biol. 2022, 71, 558–570. [Google Scholar] [CrossRef]
- Schulze, R.E. South African Atlas of Agrohydrology and Climatology. 2001. Available online: http://fred.csir.co.za/project/tmg/agrohydrology_atlas/atlas_toc.htm (accessed on 29 January 2023).
- Masters, J.C.; Génin, F.; Zhang, Y.; Pellen, R.; Huck, T.; Mazza, P.P.A.; Rabineau, M.; Doucouré, M.; Aslanian, D. Biogeographic mechanisms involved in the colonization of Madagascar by African vertebrates: Rifting, rafting and runways. J. Biogeogr. 2021, 48, 492–510. [Google Scholar] [CrossRef]
- Linder, H.P.; Verboom, G.A. The Evolution of Regional Species Richness: The History of the Southern African Flora. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 393–412. [Google Scholar] [CrossRef]
- Chorowicz, J. The East African rift system. J. Afr. Earth Sci. 2005, 43, 379–410. [Google Scholar] [CrossRef]
- Zachos, J.; Dickens, G.; Zeebe, R. An early Cenozoic perspective on greenhouse warming and carbon cycle dynamics. Nature 2008, 451, 279–283. [Google Scholar] [CrossRef] [Green Version]
- Kandziora, M.; Kadereit, J.W.; Gehrke, B. Dual colonization of the Palaearctic from different regions in the Afrotropics by Senecio. J. Biogeogr. 2017, 44, 147–157. [Google Scholar] [CrossRef]
- Assefa, A.; Ehrich, D.; Taberlet, P.; Nemomissa, S.; Brochmann, C. Pleistocene colonization of afro-alpine “sky islands” by the arctic-alpine Arabis alpina. Heredity 2007, 99, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Carlsen, T.; Bleeker, W.; Hurka, H.; Elver, R.; Brochmann, C. Biogeography and phylogeny of Cardamine (Brassicaeae). Ann. Missouri Bot. Gard. 2009, 96, 215–236. [Google Scholar] [CrossRef]
- Gehrke, B.; Kandziora, M.; Pirie, M.D. The evolution of dwarf shrubs in alpine environments: A case study of Alchemilla in Africa. Ann. Bot. 2016, 117, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Gizaw, A.; Brochmann, C.; Nemomissa, S.; Wondimu, T.; Masao, C.A.; Tusiime, F.M.; Abdi, A.A.; Oxelman, B.; Popp, M.; Dimitrov, D. Colonization and diversification in the African ‘sky islands’: Insights from fossil-calibrated molecular dating of Lychnis (Caryophyllaceae). New Phytol. 2016, 211, 719–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tusiime, F.M.; Gizaw, A.; Wondimu, T.; Masao, C.A.; Abdi, A.A.; Muwanika, V.; Trávnócek, P.; Nemonissa, S.; Popp, M.; Eilu, G.; et al. Sweet vernal grasses (Anthoxanthum) colonized African mountains along two fronts in the Late Pliocene, followed by secondary contract, polyploidization and local extinction in the Pleistocene. Mol. Ecol. 2017, 26, 3513–3532. [Google Scholar] [CrossRef] [Green Version]
- Pirie, M.D.; Kandziora, M.; Nürk, N.M.; Le Maitre, N.C.; Mugrabi de Kuppler, A.; Gehrke, B.; Oliver, E.G.H.; Bellstedt, D.U. Leaps and bounds: Geographical and ecological distance constrained the colonization of the Afromontemperate by Erica. BMC Evol. Biol. 2019, 19, 222. [Google Scholar] [CrossRef] [Green Version]
- Axelrod, D.I.; Raven, P.H. Late Cretaceous and Tertiary vegetation history of Africa. In Biogeography and Ecology of Southern Africa; Werger, M.J.A., Ed.; Springer: The Hague, The Netherlands, 1978; pp. 77–139. [Google Scholar] [CrossRef]
- Harmsen, R.; Spence, J.R.; Mahaney, W.C. Glacial interglacial cycles and development of the Afroalpine ecosystem on East-African mountains, II. Origins and development of the biotic component. J. Afr. Earth Sci. 1991, 12, 512–523. [Google Scholar] [CrossRef]
- Nathan, R.; Schurr, F.M.; Spiegel, O.; Steinitz, O.; Trakhtenbrot, A.; Tsoar, A. Mechanisms of long-distance seed dispersal. Trends Ecol. Evol. 2008, 23, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Bergh, N.G.M.; Linder, H.P. Cape diversification and repeated out-of-southern-Africa dispersal in paper daisies (Asteraceae- Gnaphalieae). Mol. Phylogenet. Evol. 2009, 51, 5–18. [Google Scholar] [CrossRef]
- Gizaw, A.; Kedebe, M.; Nemomissa, S.; Ehrich, D.; Bekele, B.; Mirré, V.; Popp, M.; Brochmann, C. Phylogeography of the heathers Erica arborea and E. trimera in the afro-alpine ‘sky islands’ inferred from AFLPs and plastid DNA sequences. Flora: Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 453–463. [Google Scholar] [CrossRef]
- Tusiime, F.M.; Gizaw, A.; Gussarova, G.; Nemomissa, S.; Popp, M.; Masao, C.A.; Wondimu, T.; Abdi, A.A.; Mirré, V.; Muwanika, V.; et al. Afro-alpine flagships revisited: Parallel adaptation, intermountain admixture and shallow genetic structuring in the giant senecios (Dendrosenecio). PLoS ONE 2020, 15, e0228979. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode | Type | Mean (sd) | % of Biogeographic Events |
|---|---|---|---|
| Within-area speciation | Sympatry | 250.2 (3.95) | 62.8 |
| Subset * | 23.38 (5.2) | 5.9 | |
| Dispersal | Founder events | 49.9 (4.28) | 12.5 |
| Range expansions | 63.18 (2.64) | 15.9 | |
| Vicariance | Vicariance | 11.56 (2.85) | 2.9 |
| Total | 398.2 (2.64) | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco-Gavaldà, C.; Galbany-Casals, M.; Susanna, A.; Andrés-Sánchez, S.; Bayer, R.J.; Brochmann, C.; Cron, G.V.; Bergh, N.G.; Garcia-Jacas, N.; Gizaw, A.; et al. Repeatedly Northwards and Upwards: Southern African Grasslands Fuel the Colonization of the African Sky Islands in Helichrysum (Compositae). Plants 2023, 12, 2213. https://doi.org/10.3390/plants12112213
Blanco-Gavaldà C, Galbany-Casals M, Susanna A, Andrés-Sánchez S, Bayer RJ, Brochmann C, Cron GV, Bergh NG, Garcia-Jacas N, Gizaw A, et al. Repeatedly Northwards and Upwards: Southern African Grasslands Fuel the Colonization of the African Sky Islands in Helichrysum (Compositae). Plants. 2023; 12(11):2213. https://doi.org/10.3390/plants12112213
Chicago/Turabian StyleBlanco-Gavaldà, Carme, Mercè Galbany-Casals, Alfonso Susanna, Santiago Andrés-Sánchez, Randall J. Bayer, Christian Brochmann, Glynis V. Cron, Nicola G. Bergh, Núria Garcia-Jacas, Abel Gizaw, and et al. 2023. "Repeatedly Northwards and Upwards: Southern African Grasslands Fuel the Colonization of the African Sky Islands in Helichrysum (Compositae)" Plants 12, no. 11: 2213. https://doi.org/10.3390/plants12112213