
Composition and Biological Activity of the Essential Oils from Wild Horsemint, Yarrow, and Yampah from Subalpine Meadows in Southwestern Montana: Immunomodulatory Activity of Dillapiole

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Plant Material
2.2. Essential Oil Composition
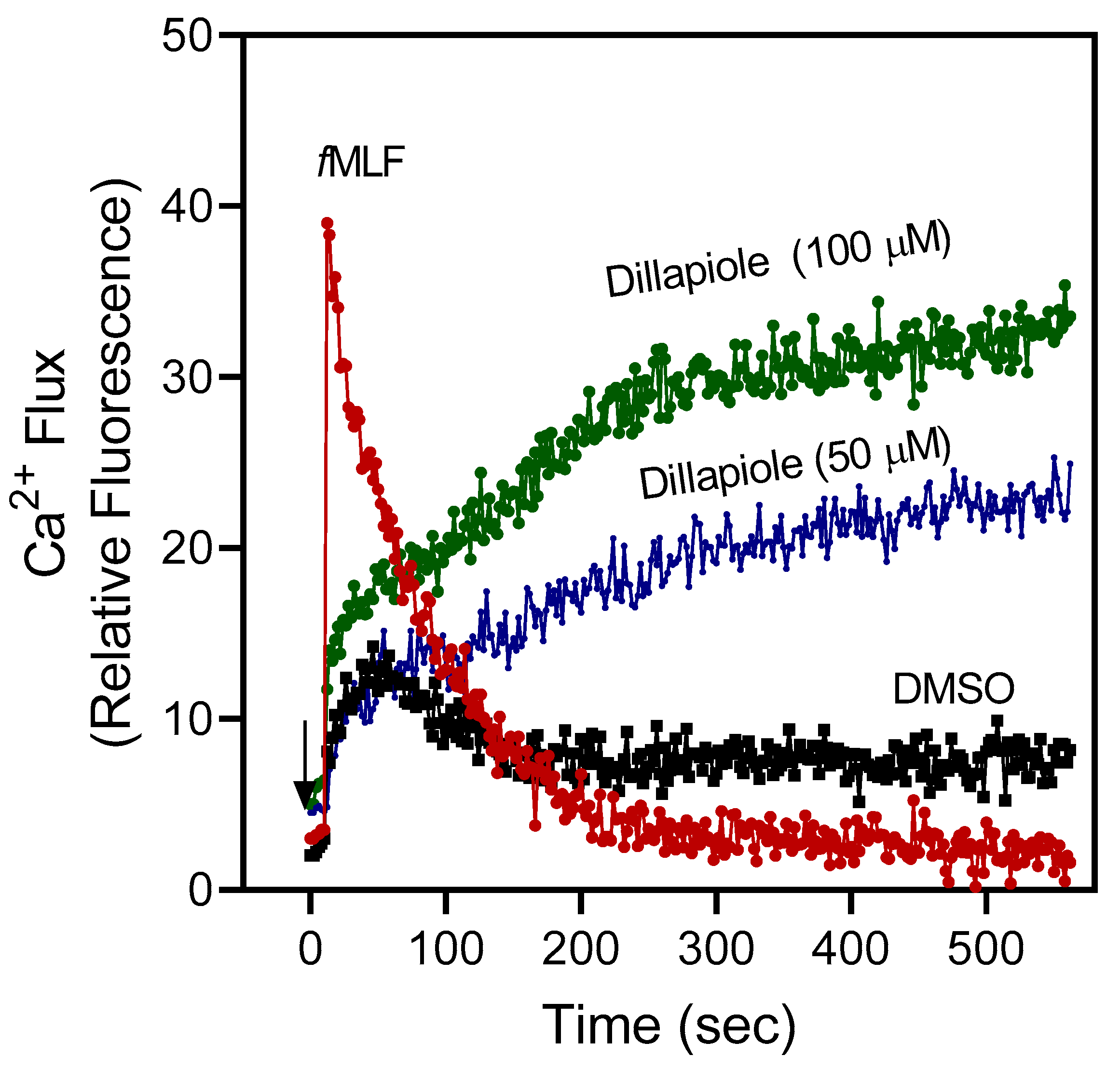
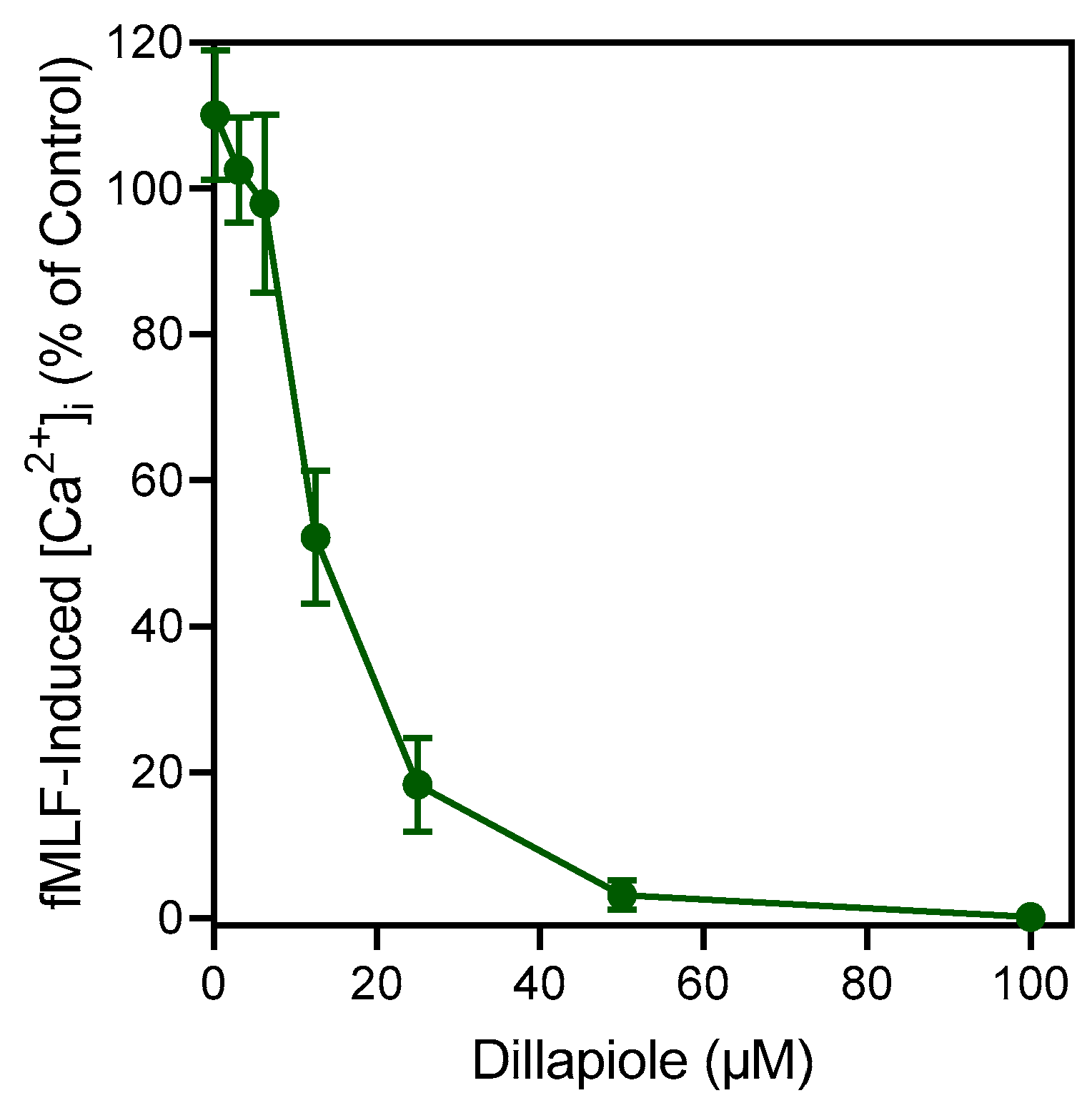
2.3. Effect of Essential Oils and Selected Component Compounds on Neutrophil Ca2+ Influx
2.4. Effect of PGI Essential Oil and Dillapiole on Neutrophil Chemotaxis
3. Materials and Methods
3.1. Materials
3.2. Essential Oil Extraction
3.3. Gas Chromatography (GC-FID) and Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
3.4. Sample Preparation for Biological Studies
3.5. Human Neutrophil Isolation
3.6. Ca2+ Mobilization Assay
3.7. Chemotaxis Assay
3.8. Cytotoxicity Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moerman, D.E. Native American Medicinal Plants: An Ethnobotanical Dictionary; Timber Press: Portland, OR, USA, 2009; p. 799. [Google Scholar]
- Chandler, R.F.; Hooper, S.N.; Harvey, M.J. Ethnobotany and phytochemistry of yarrow, Achillea millefolium, Compositae. Econ. Bot. 1982, 36, 203–223. [Google Scholar] [CrossRef]
- Ali, S.I.; Gopalakrishnan, B.; Venkatesalu, V. Pharmacognosy, phytochemistry and pharmacological properties of Achillea millefolium L.: A review. Phytother. Res. 2017, 31, 1140–1161. [Google Scholar] [CrossRef]
- Borrelli, F.; Romano, B.; Fasolino, I.; Tagliatatela-Scafati, O.; Aprea, G.; Capasso, R.; Capasso, F.; Bottazzi, E.C.; Izzo, A.A. Prokinetic effect of a standardized yarrow (Achillea millefolium) extract and its constituent choline: Studies in the mouse and human stomach. Neurogastroenterol. Motil. 2012, 24, E164–E190. [Google Scholar] [CrossRef] [PubMed]
- Sandner, G.; Heckmann, M.; Weghuber, J. Immunomodulatory activities of selected essential oils. Biomolecules 2020, 10, 1139. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, G.R.; Vasconcelos, A.B.S.; Haran, G.H.; Calisto, V.; Jothi, G.; Quintans, J.S.S.; Cuevas, L.E.; Narain, N.; Junior, L.J.Q.; Cipolotti, R.; et al. Essential oils and its bioactive compounds modulating cytokines: A systematic review on anti-asthmatic and immunomodulatory properties. Phytomedicine 2020, 73, 152854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdivieso-Ugarte, M.; Gomez-Llorente, C.; Plaza-Diaz, J.; Gil, A. Antimicrobial, antioxidant, and immunomodulatory properties of essential oils: A systematic review. Nutrients 2019, 11, 2786. [Google Scholar] [CrossRef] [Green Version]
- Beutler, B. Innate immunity: An overview. Mol. Immunol. 2004, 40, 845–859. [Google Scholar] [CrossRef]
- Bokoch, G.M. Chemoattractant signaling and leukocyte activation. Blood 1995, 86, 1649–1660. [Google Scholar] [CrossRef] [Green Version]
- Schepetkin, I.A.; Ozek, G.; Ozek, T.; Kirpotina, L.N.; Kokorina, P.I.; Khlebnikov, A.I.; Quinn, M.T. Neutrophil Immunomodulatory activity of nerolidol, a major component of essential oils from Populus balsamifera buds and propolis. Plants 2022, 11, 3399. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Ozek, G.; Ozek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. Neutrophil immunomodulatory activity of (−)-borneol, a major component of essential oils extracted from Grindelia squarrosa. Molecules 2022, 27, 4897. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Ozek, G.; Ozek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. Chemical composition and immunomodulatory activity of essential oils from Rhododendron albiflorum. Molecules 2021, 26, 3652. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Ozek, G.; Ozek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Klein, R.A.; Quinn, M.T. Neutrophil immunomodulatory activity of farnesene, a component of Artemisia dracunculus essential oils. Pharmaceuticals 2022, 15, 642. [Google Scholar] [CrossRef] [PubMed]
- Ozek, G.; Schepetkin, I.A.; Yermagambetova, M.; Ozek, T.; Kirpotina, L.N.; Almerekova, S.S.; Abugalieva, S.I.; Khlebnikov, A.I.; Quinn, M.T. Innate immunomodulatory activity of cedrol, a component of essential oils isolated from Juniperus species. Molecules 2021, 26, 7644. [Google Scholar] [CrossRef]
- Tadic, V.; Arsic, I.; Zvezdanovic, J.; Zugic, A.; Cvetkovic, D.; Pavkov, S. The estimation of the traditionally used yarrow (Achillea millefolium L. Asteraceae) oil extracts with anti-inflamatory potential in topical application. J. Ethnopharmacol. 2017, 199, 138–148. [Google Scholar] [CrossRef]
- Bogdan, A.; Colţun, M. Agastache urticifolia (Benth.) Kuntze–aromatic plant introduced and researched in the Botanical Garden. Rev. Bot. 2020, 21, 158–160. [Google Scholar] [CrossRef]
- Wilson, T.M.; Davis, A.; Sonstrom, R.E.; Neill, J.L.; Ziebarth, E.A.; Poulson, A.; Carlson, R.E. Essential oil composition and enantioselective profile of Agastache urticifolia (Lamiaceae) and Monardella odoratissima (Lamiaceae) from Utah. Molecules 2023, 28, 2249. [Google Scholar] [CrossRef]
- Zielinska, S.; Matkowski, A. Phytochemistry and bioactivity of aromatic and medicinal plants from the genus Agastache (Lamiaceae). Phytochem. Rev. 2014, 13, 391–416. [Google Scholar] [CrossRef] [Green Version]
- Bogdan, A.; Colţun, M.; Gille, E.; Necula, R.D.; Grigoraş, V. The biology and the chemical composition of the essential oil of the specie Agastache urticifolia (Benth). Kuntze. In Proceedings of the V International Scientific and Practical Conference within the Framework of the VI Scientific Forum “Science Week in Kruty”, Kruty, Ukraine, 11 March 2021; pp. 38–44. [Google Scholar]
- Myadelets, M.A.; Vorobyeva, T.A.; Domrachev, D.V. Composition of the essential oils of some species belonging to genus Agastache clayton ex Gronov (Lamiaceae) cultivated under the conditions of the Middle Ural. Chem. Sustain. Dev. 2013, 21, 397–401. [Google Scholar]
- Viljoen, A.M.; Moolla, A.; van Vuuren, S.F.; van Zyl, R.L.; Baser, K.H.C.; Demirci, B.; Ozek, T.; Trinder-Smith, T.H. The biological activity and essential oil composition of 17 Agathosma (Rutaceae) species. J. Essent. Oil Res. 2006, 18, 2–16. [Google Scholar] [CrossRef]
- Alan, S.; Kurkcuoglu, M.; Baser, K.H.C. Composition of essential oils of Calamintha nepeta (L.) Savi Subsp nepeta and Calamintha nepeta (L.) Savi Subsp glandulosa (Req.) PW Ball. Asian J. Chem. 2011, 23, 2357–2360. [Google Scholar]
- Kaiser, R.; Lamparsky, D.; Schudel, P. Analysis of Buchu leaf oil. J. Agr. Food Chem. 1975, 23, 943–950. [Google Scholar] [CrossRef]
- Orav, A.; Arak, E.; Raal, A. Phytochemical analysis of the essential oil of Achillea millefolium L. from various European countries. Nat. Prod. Res. 2006, 20, 1082–1088. [Google Scholar] [CrossRef]
- Falconieri, D.; Piras, A.; Porcedda, S.; Marongiu, B.; Goncalves, M.J.; Cabral, C.; Cavaleiro, C.; Salgueiro, L. Chemical composition and biological activity of the volatile extracts of Achillea millefolium. Nat Prod Commun 2011, 6, 1527–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tampe, J.; Parra, L.; Huaiquil, K.; Mutis, A.; Quiroz, A. Repellent effect and metabolite volatile profile of the essential oil of Achillea millefolium against Aegorhinus nodipennis (Hope) (Coleoptera: Curculionidae). Neotrop. Entomol. 2015, 44, 279–285. [Google Scholar] [CrossRef]
- Toplan, G.G.; Taskin, T.; Iscan, G.; Goger, F.; Kurkcuoglu, M.; Civas, A.; Ecevit-Genc, G.; Mat, A.; Baser, K.H.C. Comparative studies on essential oil and phenolic content with in vitro antioxidant, anticholinesterase, antimicrobial activities of Achillea biebersteinii Afan. and A. millefolium subsp. millefolium Afan. L. growing in Eastern Turkey. Molecules 2022, 27, 1956. [Google Scholar] [CrossRef]
- Acimovic, M.; Zoric, M.; Zheljazkov, V.D.; Pezo, L.; Cabarkapa, I.; Jeremic, J.S.; Cvetkovic, M. Chemical characterization and antibacterial activity of essential oil of medicinal plants from Eastern Serbia. Molecules 2020, 25, 5482. [Google Scholar] [CrossRef]
- El-Kalamouni, C.; Venskutonis, P.R.; Zebib, B.; Merah, O.; Raynaud, C.; Talou, T. Antioxidant and antimicrobial activities of the essential oil of Achillea millefolium L. grown in France. Medicines 2017, 4, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millefolii Herba, Y. European Pharmacopoeia. European Directorate for the Quality of Medicines & HealthCare of the Council of Europe (EDQM), 9th ed.; Council of Europe: Strasbourg, France, 2016; Volume 1, pp. 1564–1565. [Google Scholar]
- Raal, A.; Orav, A.; Arak, E. Essential oil content and composition in commercial Achillea millefolium L. herbs from different countries. J. Essent. Oil Bear. Plants 2012, 15, 22–31. [Google Scholar] [CrossRef]
- Rauchensteiner, F.; Nejati, S.; Saukel, J. The Achillea millefolium group (Asteraceae) in Middle Europe and the Balkans: A diverse source for the crude drug Herba Millefolii. J. Trad. Medi. 2004, 21, 113–119. [Google Scholar]
- Schepetkin, I.A.; Ozek, G.; Ozek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. Chemical composition and immunomodulatory activity of Hypericum perforatum essential oils. Biomolecules 2020, 10, 916. [Google Scholar] [CrossRef] [PubMed]
- Naya, Y.; Nagahama, Y.; Kotake, M. Volatile components of Ledum palustre Var Nipponicum Et Yesoense. Heterocycles 1978, 10, 29–36. [Google Scholar] [CrossRef]
- Madyastha, K.M.; Raj, C.P. Stereoselective hydroxylation of 4-methyl-2-cyclohexenone in rats: Its relevance to R-(+)-pulegone-mediated hepatotoxicity. Biochem. Biophys. Res. Comm. 2002, 297, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Kushnarenko, S.V.; Ozek, G.; Kirpotina, L.N.; Sinharoy, P.; Utegenova, G.A.; Abidkulova, K.T.; Ozek, T.; Baser, K.H.; Kovrizhina, A.R.; et al. Modulation of Human neutrophil responses by the essential oils from Ferula akitschkensis and their constituents. J. Agric. Food Chem. 2016, 64, 7156–7170. [Google Scholar] [CrossRef] [Green Version]
- Schepetkin, I.A.; Kushnarenko, S.V.; Ozek, G.; Kirpotina, L.N.; Utegenova, G.A.; Kotukhov, Y.A.; Danilova, A.N.; Ozek, T.; Baser, K.H.; Quinn, M.T. Inhibition of human neutrophil responses by the essential oil of Artemisia kotuchovii and its constituents. J. Agric. Food Chem. 2015, 63, 4999–5007. [Google Scholar] [CrossRef] [Green Version]
- Kreutz, T.; Carneiro, S.B.; Soares, K.D.; Limberger, R.P.; Apel, M.A.; Veiga, V.F.; Koester, L.S. Aniba canelilla (Kunth) Mez essential oil-loaded nanoemulsion: Improved stability of the main constituents and in vitro antichemotactic activity. Ind. Crops Prod. 2021, 171, 113949. [Google Scholar] [CrossRef]
- Maciel, A.J.; Lacerda, C.P.; Danielli, L.J.; Bordignon, S.A.L.; Fuentefria, A.M.; Apel, M.A. Antichemotactic and antifungal action of the essential oils from Cryptocarya aschersoniana, Schinus terebinthifolia, and Cinnamomum amoenum. Chem. Biodiver. 2019, 16, 204. [Google Scholar] [CrossRef] [PubMed]
- Soares, K.D.; Bordignon, S.A.L.; Apel, M.A. Chemical composition and anti-inflammatory activity of the essential oils of Piper gaudichaudianum and Piper mikanianum. J. Ethnopharmacol. 2022, 297, 115533. [Google Scholar] [CrossRef] [PubMed]
- Santana, A.I.; Vila, R.; Canigueral, S.; Gupta, M.P. Chemical composition and biological activity of essential oils from different species of Piper from Panama. Planta Med. 2016, 82, 986–991. [Google Scholar] [CrossRef]
- Ferreira, O.O.; Cruz, J.N.; de Moraes, A.A.B.; Franco, C.D.P.; Lima, R.R.; Dos Anjos, T.O.; Siqueira, G.M.; do Nascimento, L.D.; Cascaes, M.M.; de Oliveira, M.S.; et al. Essential oil of the plants growing in the Brazilian Amazon: Chemical composition, antioxidants, and biological applications. Molecules 2022, 27, 4373. [Google Scholar] [CrossRef]
- Vila, R.; Tomi, M.; Mundina, M.; Santana, A.I.; Solis, P.N.; Arce, J.B.L.; Iclina, J.L.B.; Iglesias, J.; Gupta, M.P.; Casanova, J.; et al. Unusual composition of the essential oils from the leaves of Piper aduncum. Flavour Fragr. J. 2005, 20, 67–69. [Google Scholar] [CrossRef]
- Guetat, A.; Abdelwahab, A.T.; Yahia, Y.; Rhimi, W.; Alzahrani, A.K.; Boulila, A.; Cafarchia, C.; Boussaid, M. Deverra triradiata Hochst. ex Boiss. from the Northern Region of Saudi Arabia: Essential oil profiling, plant extracts and biological activities. Plants 2022, 11, 1543. [Google Scholar] [CrossRef] [PubMed]
- Brazao, M.A.B.; Brazao, F.V.; Maia, J.G.S.; Monteiro, M.C. Antibacterial activity of the Piper aduncum oil and dillapiole, its main constituent, against multidrug-resistant strains. Bol. Latinoam. Caribe Plantas 2014, 13, 517–526. [Google Scholar]
- de Almeida, R.R.P.; Souto, R.N.P.; Bastos, C.N.; da Silva, M.H.L.; Maia, J.G.S. Chemical variation in Piper aduncum and biological properties of its dillapiole-rich essential oil. Chem. Biodiver. 2009, 6, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Parise, R.; Pasqualoto, K.F.M.; Magri, F.M.M.; Ferreira, A.K.; da Silva, B.A.V.G.; Damiao, M.C.F.C.B.; Tavares, M.T.; Azevedo, R.A.; Auada, A.V.V.; Polli, M.C.; et al. Dillapiole as antileishmanial agent: Discovery, cytotoxic activity and preliminary SAR studies of dillapiole analogues. Arch. Pharm. 2012, 345, 934–944. [Google Scholar] [CrossRef]
- Rojas-Martinez, R.; Arrieta, J.; Cruz-Antonio, L.; Arrieta-Baez, D.; Velazquez-Mendez, A.M.; Sanchez-Mendoza, M.E. Dillapiole, isolated from Peperomia pellucida, shows gastroprotector activity against ethanol-induced gastric lesions in Wistar rats. Molecules 2013, 18, 11327–11337. [Google Scholar] [CrossRef] [Green Version]
- Parise, R.; Pastrello, M.; Camerlingo, C.E.P.; Silva, G.J.; Agostinho, L.A.; de Souza, T.; Magri, F.M.M.; Ribeiro, R.R.; Brandt, C.A.; Polli, M.C. The anti-inflammatory activity of dillapiole and some semisynthetic analogues. Pharmaceut. Biol. 2011, 49, 1173–1179. [Google Scholar] [CrossRef]
- Ferreira, A.K.; de-Sa, P.L.; Pasqualoto, K.F.M.; de Azevedo, R.A.; Camara, D.A.D.; Costa, A.S.; Figueiredo, C.R.; Matsuo, A.L.; Massaoka, M.H.; Auada, A.V.V.; et al. Cytotoxic effects of dillapiole on MDA-MB-231 cells involve the induction of apoptosis through the mitochondrial pathway by inducing an oxidative stress while altering the cytoskeleton network. Biochimie 2014, 99, 195–207. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, S.B.; Kreutz, T.; Limberger, R.P.; Teixeira, H.F.; da Veiga Júnior, V.F.; Koester, L.S. Piper aduncum essential oil rich in dillapiole: Development of hydrogel-thickened nanoemulsion and nanostructured lipid carrier intended for skin delivery. Pharmaceutics 2022, 14, 2525. [Google Scholar] [CrossRef]
- Ozek, G.; Ishmuratova, M.; Tabanca, N.; Radwan, M.M.; Goger, F.; Ozek, T.; Wedge, D.E.; Becnel, J.J.; Cutler, S.J.; Can Baser, K.H. One-step multiple component isolation from the oil of Crinitaria tatarica (Less.) Sojak by preparative capillary gas chromatography with characterization by spectroscopic and spectrometric techniques and evaluation of biological activity. J. Sep. Sci. 2012, 35, 650–660. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. High-throughput screening for small-molecule activators of neutrophils: Identification of novel N-formyl peptide receptor agonists. Mol. Pharmacol. 2007, 71, 1061–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Location | Latitude (N) | Longitude (E) | Altitude (m) | Plant Material | Date of Collection | Yield (%) |
|---|---|---|---|---|---|---|---|
| A. urticifolia | Hyalite Canyon, Bozeman, MT, USA | 45.48990° | 111.00091° | 2272 | leaves/flowers | 07/2021 | 0.2/0.5 |
| A. millefolium | Hyalite Canyon, Bozeman, MT, USA | 45.48346° | 110.97882° | 2042 | inflorescence | 07/2021 | 0.2 |
| P. gairdneri | Hyalite Canyon, Bozeman, MT, USA | 45.49671° | 110.98859° | 1978 | inflorescence | 07/2021 | 1.9 |
| RRI | Compound | AUF | AUL | AMI | PGI | RRI | Compound | AUF | AUL | AMI | PGI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1032 | α-Pinene | t | t | 1.2 | 0.2 | 1704 | γ-Muurolene | t | |||
| 1035 | α-Thujene | t | t | t | t | 1706 | α-Terpineol | t | 1.9 | 1.1 | |
| 1048 | 2-Methyl-3-buten-2-ol | t | t | 1715 | γ-Terpineol | t | |||||
| 1076 | Camphene | 2.3 | t | 1719 | Borneol | 1.2 | |||||
| 1093 | Hexanal | t | t | 1720 | trans-Sabinol | 0.2 | |||||
| 1118 | β-Pinene | t | t | 1.9 | t | 1726 | Germacrene D | 0.1 | 0.1 | ||
| 1132 | Sabinene | 0.1 | 0.1 | 7.0 | 0.1 | 1748 | Piperitone | 0.1 | 0.7 | ||
| 1136 | Isoamyl acetate | t | 0.4 | 1751 | Carvone | 0.1 | 0.2 | t | |||
| 1174 | Myrcene | 0.4 | 0.4 | t | 0.2 | 1773 | δ-Cadinene | 0.2 | |||
| 1176 | α-Phellandrene | t | 0.1 | 1797 | p-Methyl acetophenone | t | 0.6 | ||||
| 1185 | Isobutyl 2-methyl butyrate | t | t | 1802 | Cumin aldehyde | 0.2 | |||||
| 1188 | α-Terpinene | 0.2 | t | 1811 | p-Mentha-1,3-dien-7-al | 0.4 | |||||
| 1195 | Dehydro-1,8-cineole | t | 0.2 | 1814 | p-Mentha-1,5-dien-7-ol | 0.7 | |||||
| 1203 | 2-Methyl butyl isobutyrate | t | 1838 | 2-Phenylethyl acetate | 0.1 | 0.2 | |||||
| 1203 | Limonene | 4.2 | 6.2 | 0.2 | 1.3 | 1845 | trans-Carveol | t | |||
| 1213 | 1,8-Cineole | t | t | 17.0 | 1849 | Pulegone epoxide | 0.4 | ||||
| 1218 | β-Phellandrene | 0.1 | 0.1 | 0.5 | 1864 | p-Cymen-8-ol | t | 14.1 | |||
| 1225 | (Z)-3-Hexenal | t | 0.2 | 1865 | Isopiperitenone | 0.5 | 0.1 | ||||
| 1244 | Amyl furan | t | 1877 | TMMT | 1.1 | 0.8 | |||||
| 1246 | (Z)-β-Ocimene | 0.1 | 0.1 | t | 0.4 | 1894 | CMMT | 0.5 | 0.4 | ||
| 1255 | γ-Terpinene | t | 0.4 | 2.4 | 1898 | 1,11-Oxidocalamenene | 0.3 | ||||
| 1266 | (E)-β-Ocimene | 0.5 | 0.3 | 0.3 | 1949 | Piperitenone | 1.6 | 0.8 | |||
| 1266 | 3-Octanone | t | 1969 | cis-Jasmone | 0.2 | ||||||
| 1280 | p-Cymene | t | 0.7 | 1.1 | 1998 | 8,9-dehydrothymol | 0.1 | t | |||
| 1285 | Isoamyl isovalerate | t | t | 2008 | Caryophyllene oxide | 0.3 | 0.4 | 0.5 | |||
| 1286 | MBMB | 0.1 | 2016 | Isoamyl phenyl acetate | 0.1 | ||||||
| 1290 | Terpinolene | t | 0.1 | 12.0 | 2045 | Carotol | 0.2 | ||||
| 1384 | α-Pinene oxide | 0.1 | 2050 | (E)-Nerolidol | 0.2 | ||||||
| 1386 | Octenyl acetate | t | 2068 | Hexahydro-farnesyl acetone | 0.1 | ||||||
| 1400 | Nonanal | t | 2074 | Caryophylla-2(12),6(13)-dien-5-one | 0.3 | ||||||
| 1408 | 1,3,8-p-Menthatriene | 0.1 | 2094 | p-Cresol | 0.2 | ||||||
| 1413 | Rose furan | t | 2096 | Elemol | 1.7 | ||||||
| 1437 | α-Thujone | 17.1 | 0.3 | 2096 | (E)-Methyl cinnamate | 0.3 | |||||
| 1443 | 2,5- Dimethylstyrene | t | 2100 | Heneicosane | 0.3 | ||||||
| 1451 | β-Thujone | 0.1 | 14.9 | 2103 | Guaiol | 0.1 | |||||
| 1452 | α,p-Dimethylstyrene | t | 0.6 | 2113 | Cumin alcohol | 0.3 | |||||
| 1452 | 1-Octen-3-ol | t | 2115 | 4-Hydroxy-4-methyl-cyclohex-2-enone | 6.2 | ||||||
| 1458 | cis-1,2-Limonene epoxide | t | 2144 | Spathulenol | 0.1 | 0.2 | |||||
| 1474 | trans-Sabinene hydrate | 0.3 | 2181 | Isothymol | 0.3 | ||||||
| 1475 | Menthone | 2.7 | 25.7 | 2183 | γ-Decalactone | 0.6 | |||||
| 1497 | α-Copaene | 0.2 | 2184 | cis-p-Menth-3-en-1,2-diol | 0.4 | ||||||
| 1497 | Menthofuran | 0.1 | 2185 | γ-Eudesmol | 0.9 | ||||||
| 1503 | Isomenthone | 2.6 | 29.1 | 2192 | Nonanoic acid | 0.1 | |||||
| 1532 | Camphor | 13.0 | 2195 | Fokienol | 0.1 | ||||||
| 1541 | Benzaldehyde | 0.2 | 0.4 | 2209 | T-Muurolol | t | |||||
| 1553 | Linalool | 0.3 | 2221 | Isocarvacrol | 0.3 | ||||||
| 1556 | cis-Sabinene hydrate | 0.2 | 2228 | Eremoligenol | 0.1 | ||||||
| 1571 | trans-p-Menth-2-en-1-ol | 0.1 | 2245 | Elemicine | 1.2 | ||||||
| 1583 | cis-Isopulegone | 0.5 | 0.2 | 2250 | α-Eudesmol | 0.5 | |||||
| 1590 | Bornyl acetate | 1.7 | 2250 | Fukinanolide | 0.6 | ||||||
| 1598 | trans-Isopulegone | 0.5 | 2255 | α-Cadinol | t | ||||||
| 1611 | Terpinen-4-ol | 2.5 | 0.4 | 2257 | β-Eudesmol | 1.1 | |||||
| 1612 | β-Caryophyllene | 0.5 | 0.3 | 0.3 | 0.1 | 2272 | Copaborneol | t | |||
| 1618 | Camphene hydrate | 0.2 | 2290 | Guaia-3,9-dien-11-ol | 3.2 | ||||||
| 1626 | MMO | t | 2296 | Myristicine | 0.1 | ||||||
| 1638 | cis-p-Menth-2-en-1-ol | 0.1 | 2303 | Menthofurolactone | 0.2 | t | |||||
| 1639 | trans-p-Mentha-2,8-dien-1-ol | 0.5 | 0.6 | 2316 | Caryophylladienol I | 0.6 | |||||
| 1642 | Thuj-3-en10-al | 0.2 | 2324 | Caryophylladienol II | 0.2 | ||||||
| 1648 | Myrtenal | 0.6 | 2368 | Eudesma-4(15),7-diene-1-β-ol | t | ||||||
| 1651 | Sabinaketone | 0.1 | 2384 | Dillapiole | 30.3 | ||||||
| 1658 | Sabinyl acetate | 0.1 | 0.2 | 2420 | 2-Methyl isoborneol * | 1.1 | |||||
| 1662 | Pulegone | 78.9 | 28.8 | 0.1 | 2622 | Phytol | t | ||||
| 1678 | cis-p-Mentha-2,8-dien-1-ol | 0.5 | 2655 | Benzyl benzoate | 1.9 | ||||||
| 1682 | δ-Terpineol | 0.6 | 2758 | Artedouglasia oxide B | 0.6 | ||||||
| 1690 | Cryptone | 0.1 | 0.3 | 0.2 | |||||||
| Compounds | AUF | AUL | AMI | PGI |
|---|---|---|---|---|
| Monoterpene hydrocarbons | 5.4 | 7.2 | 14.0 | 19.3 |
| Oxygenated monoterpenes | 90.6 | 88.7 | 73.6 | 18.4 |
| Sesquiterpene hydrocarbons | 0.6 | 0.3 | 0.8 | 0.1 |
| Oxygenated sesquiterpenes | 0.4 | 0.6 | 9.8 | 1.4 |
| Oxygenated diterpenes | t | |||
| Phenylpropanoids | 31.6 | |||
| Substituted cyclohexanones | 6.2 | |||
| Others | 0.4 | 1.7 | 0.5 | 3.6 |
| Total | 97.4 | 98.5 | 98.2 | 80.6 |
| Essential Oil or Pure Compound | Direct Activation | Inhibition of fMLF-Induced Response a |
|---|---|---|
| EC50 (µg/mL); (Efficacy, %) | IC50 (µg/mL) | |
| AUF | 28.5 ± 2.1 (130) | 43.0 ± 2.8 |
| AUL | 43.5 ± 6.2 (150) | 25.0 ± 4.2 |
| AMI | 41.5 ± 0.7 (140) | 24.5 ± 2.1 |
| PGI | 30.6 ± 4.2 (70) | 4.3 ± 2.2 |
| EC50 (µM); (Efficacy, %) | IC50 (µM) | |
| α-Thujene | N.A. | N.A. |
| α/β-Thujone | N.A. | N.A. |
| Menthone | N.A. | N.A. |
| Isomenthone | N.A. | N.A. |
| Pulegone | N.A. | N.A. |
| Dillapiole | 19.3 ± 1.4 (65) | 13.9 ± 4.2 |
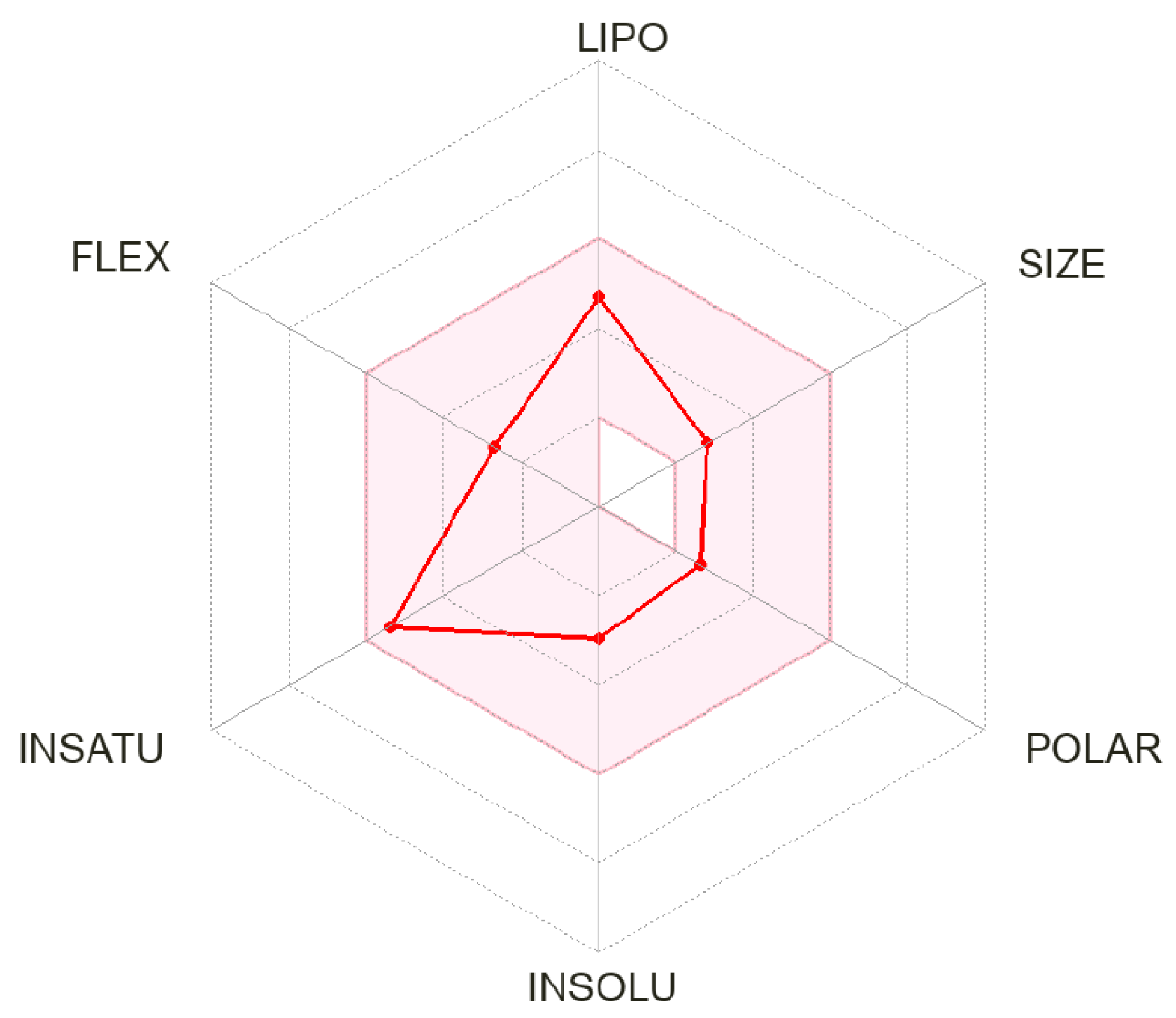
| Molecular Descriptor | Property |
|---|---|
| Formula | C12H14O4 |
| M.W. | 222.24 |
| Heavy atoms | 16 |
| Fraction Csp3 | 0.33 |
| Rotatable bonds | 4 |
| H-bond acceptors | 4 |
| H-bond donors | 0 |
| MR | 59.59 |
| tPSA | 36.92 |
| iLogP | 2.82 |
| BBB permeation | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schepetkin, I.A.; Özek, G.; Özek, T.; Kirpotina, L.N.; Klein, R.A.; Khlebnikov, A.I.; Quinn, M.T. Composition and Biological Activity of the Essential Oils from Wild Horsemint, Yarrow, and Yampah from Subalpine Meadows in Southwestern Montana: Immunomodulatory Activity of Dillapiole. Plants 2023, 12, 2643. https://doi.org/10.3390/plants12142643
Schepetkin IA, Özek G, Özek T, Kirpotina LN, Klein RA, Khlebnikov AI, Quinn MT. Composition and Biological Activity of the Essential Oils from Wild Horsemint, Yarrow, and Yampah from Subalpine Meadows in Southwestern Montana: Immunomodulatory Activity of Dillapiole. Plants. 2023; 12(14):2643. https://doi.org/10.3390/plants12142643
Chicago/Turabian StyleSchepetkin, Igor A., Gulmira Özek, Temel Özek, Liliya N. Kirpotina, Robyn A. Klein, Andrei I. Khlebnikov, and Mark T. Quinn. 2023. "Composition and Biological Activity of the Essential Oils from Wild Horsemint, Yarrow, and Yampah from Subalpine Meadows in Southwestern Montana: Immunomodulatory Activity of Dillapiole" Plants 12, no. 14: 2643. https://doi.org/10.3390/plants12142643