1. Introduction
The Asteraceae plants, the world’s largest and most diversified flowering plant family, containing 1600–1700 genera and about 25,000–35,000 species, is subdivided into 13 subfamilies, including Barnadesioideae, Cichorioideae, and Asteroideae [
1,
2]. The phylogeny and diversifications of the Asteraceae family have been impeded by the absence of extensive research into several genera and representative species. It is known that chicory and endive were grown in ancient Egypt, and currently they represent important components in Mediterranean diets. Both are now cultivated worldwide, especially in the Mediterranean basin. The subfamily Cichorioideae has the tribe Lactuceae, which contains the
Cichorium genus. This genus includes
Cichorium intybus L. (chicory) and
C. endivia L. (endive), representing two distinct species recognized according to provenance. The European flora is referred to the
C. intybus, C.
spinosum, and
C. endivia species, subdividing the latter into two subspecies; one is cultivated (subsp.
Endivia) (cultivated) and the other is wild (subsp.
divaricatum) as detailed in Tutin et al. [
3]. The reference to the Italian flora combines three wild species of
C. intybus and
C. spinosum, with the botanical variety
glabratum (Presl) Fiori, along with
C. pumilum and
C. endivia in a single cultivated species [
4]. Asteraceae in Egypt includes one wild species of
Cichorium [
5].
Seven
Cichorium species were characterized morphologically by a revision of Bedarff [
6].
C. endivia and
C. intybus split further into two subspecies; C.
endivia contained subsp.
endivia and subsp.
divaricatum, while
C. intybus was subdivided into subsp.
intybus and subsp.
glabratum. This diverges from the Flora Europaea database, Royal Botanical Gardens, which indicates only three subspecies of
C. intybus: subsp.
foliosum (Hegi) Janch., subsp.
glabratum (C. Presl) Arcang., and subsp.
sativum (Bisch.) Janch.
The integration between morphological characteristics and molecular findings characterized the two cultivated and well-known
Cichorium species,
C. intybus and
C. endivia, as well as the wild species of
C. pumilum and
C. spinosum [
7]. In addition, two other species were not found in Europe,
C. calvum Schultz-Bip and
C. bottae Defl.
C. calvum is prevalent in hot and dry areas of Southwestern Asia and the Middle East., while
C. bottae is found only in Saudi Arabia and Yemen. Finally, in Italian flora, three species of the genus were documented:
C. endivia, with the two subspecies
pumilum (Jacq) Cout. and
endivia Hegi,
C. intybus, altogether with the two subspecies
glabratum (C. Presl) Arcang., and
intybus, and
C. spinosum [
8].
Recently, Bartolucci et al. [
9] published an updated list of the vascular flora native to Italy, including the genus
Cichorium with three species:
C. endivia L. subsp.
pumilum (Jacq.) Cout.,
C. intybus L., and
C. spinosum L.
Cichorium intybus could be classified into four varieties based on the purpose and usage for which it was cultivated [
10].
C. intybus subsp.
intybus can be categorized into five major groups, including all cultivated forms of chicory [
11,
12]. Apart from wild accessions, the first group refers to the var.
foliosum, which contains the Witloof chicory. It is suggested that the Witloof chicory (
Cichorium intybus L. var.
foliosum, syn. Belgian endive, chicon) taken from Magdeburg roots should be classified under var.
sativum with all the other root types; many reviews referred to Witloof as having botanical characteristics of the variety
foliosum [
7,
12,
13,
14].
Jana and Mukherjee [
15] described the testa structure of
C. endivia and
C. intybus.
C. intybus, a biennial species, which has a characteristic inflorescence (capitulum) unique in the family containing 15–25 hermaphrodite flowers with an involucre protecting the receptacle. Every flower possesses a gamopetalous and ligulate corolla with five filamentous stamens. Stamens form a column with fused anthers surrounding the pistil with a bifid stigma [
16].
Molecular investigations have been efficiently employed to evaluate the identification and relationship of various taxa. Numerous PCR-based molecular markers are widely used such as random amplified fragment DNA (RAPD), inter-simple sequence repeats (ISSR), and start codon target (SCoT) applying in various taxa [
17,
18,
19,
20], which can be used to estimate genetic relationships and variations between and within taxa [
21,
22,
23]. A limited number of molecular studies have integrated with morphological differentiations in relation to biodiversity, identification, and taxonomy in the taxa of chicory. The genetic identification and differentiation of a taxon are applied in various species such as
Brassica [
24], broccoli and cauliflower [
25], lettuce and Jew’s mallow [
26], and tomato [
27]. In
Cichorium, various molecular markers were applied [
7,
28,
29,
30,
31,
32,
33,
34,
35,
36,
37]. Some of these markers have been used for genetic map construction [
31,
32], genetic variation [
33,
34], gene flow [
35], population structure [
37], and hybridization [
36].
The Egyptian chicory (“Sreece” according to its local Arabic name) belongs to the
C. endivia sub-species
pumilum in the Asteraceae family. However, several published articles from Egypt still incorrectly assume that the Egyptian chicory belongs to the
C. intybus species [
38,
39,
40]. The integrated approach using micro-macro morphological assessment and molecular techniques could help to improve authentication and verification of the species and taxa. With the lack of extensive sampling of several genera and representative species, the family’s phylogeny and diversifications have been hampered by the limited availability of research covering species and varietal diversity, along with the high resemblance among species.
This study highlights the morphological and molecular characteristics of five taxa of the genus Cichorium through assessment and discrimination to determine the relationships among taxa. The micro- and macro-morphological data generated a relationship among the taxa of Cichorium, while the PCR-based markers provided information on Cichorium’s genetic relatedness. The goal of this study is to present analyses to accurately classify the studied chicory species. In order to resolve the classification ambiguity, we offer a robust argument by documenting morphological and genetic differences between the C. endivia and C. intybus species of chicory.
3. Discussion
The morphological characteristics studied in this research, such as cypsela and pappus, using scanning electron micrographs, have already provided valuable information for specific genera [
41,
42]. The five taxa of
Cichorium showed morphological differences. Identification and classification between taxa depend mostly on morphological characteristics. The studied taxa have a wide range of variations in characteristics associated with leaves (shape, type, texture, margin, and apex), cypselae (shape, color, and surface pattern), and trichomes (glands).
Cichorium intybus is characterized by a spathulate leaf shape, a cypsela sulcate papillate surface pattern, a raised anticlinal wall, a concave periclinal wall, and a pappus type with scabrous scales.
Cichorium endivia is characterized by having a leaf shape, simple or pinnately lobed, a slightly raised anticlinal wall, a slightly concave periclinal wall, and a pappus type with paleaceous scales to form a crown. The examined taxa demonstrated variations in their morphological features, such as leaf morphology, cypsela, and trichome characteristics. The five taxa had stomata on both surfaces, particularly the abaxial epidermis. Although they are very similar due to their morphological characteristics,
C. intybus and
C. endivia have always been categorized into two different species, morphologically described by Kiers et al. [
43]. Kiers et al. [
7] described
C. intybus and
C. endivia, the two cultivated and most known species, using an integration of morphological features with molecular approaches. Our results are in agreement with the view of Kiers et al. [
7,
43] and Raulier et al. [
12].
Genetic identification of
Cichorium using PCR-based molecular markers (RAPD, ISSR, and SCoT) enabled the detection of polymorphism to reveal the relationships between the studied taxa. Genetic analysis is crucial for managing the entire classification and identification, as well as genetic improvement [
44]. The efficiency of a marker for discriminating species depends mainly upon the resultant polymorphism [
22,
24,
45]. Markers of ISSR and SCoT have the potential to reveal polymorphism and offer a higher capacity for the determination of intra- and inter-genomic variations than other arbitrary primers such as RAPDs [
21,
26,
46]. The difference in resolution resulting from RAPDs and ISSRs can be explained by the fact that different positions of the genome are targeted by the two-marker techniques [
21]. Therefore, the ability to reveal genetic variability among and within species is directly related to the detected percentage of polymorphism using each marker than the technique employed. Similar conclusions were obtained by Mahdy [
26] on lettuce and Jew’s mallow and Gupta et al. [
46] on Jatropha. Kiers et al. [
7] studied endive and chicory cultivars using diagnostic AFLP markers in
Cichorium species. Our results are in general agreement with several previous studies [
28,
29,
30,
31].
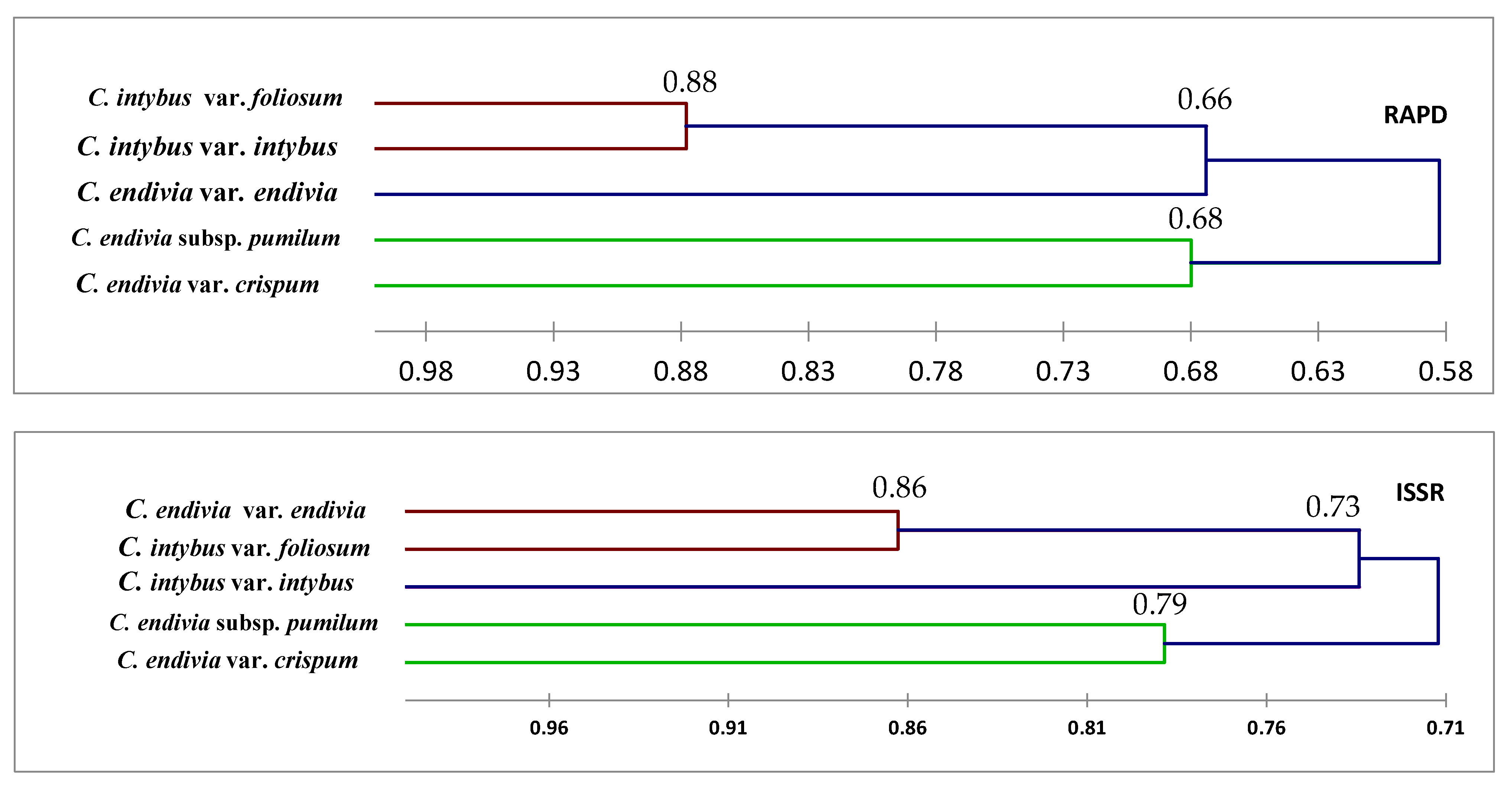
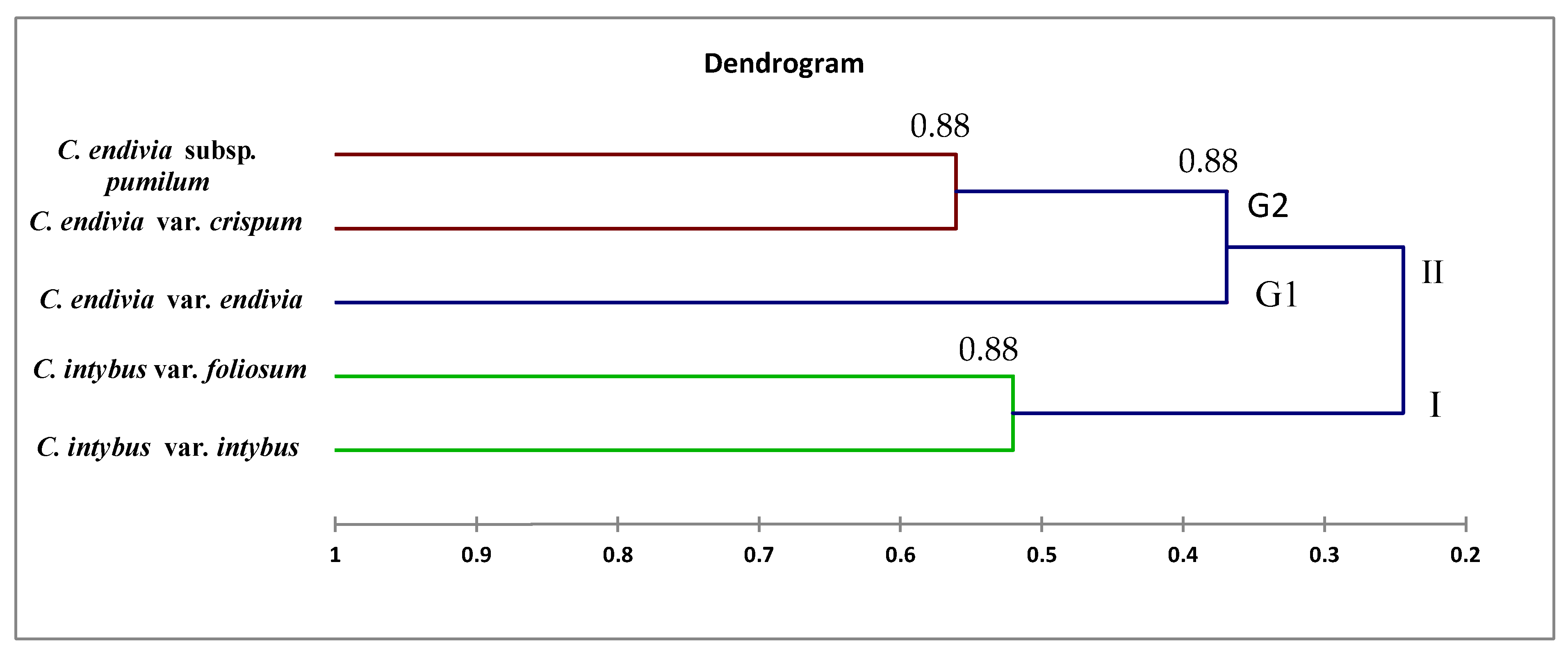
Dendrograms, as illustrated in
Figure 8,
Figure 9 and
Figure 10, have shown a correspondence between molecular markers and morphological attributes. The differences among the dendrograms generated by markers could be partially explained by the different number of PCR products analyzed, as presented in
Table 6. This highlights the importance of higher genome coverage and loci number in estimating the genetic relationships among the tested taxa. Loarce et al. [
47] obtained similar findings in barley. This could also be explained by the low reproducibility of RAPD markers [
48]. The differences in the clustering pattern of genotypes using RAPD and ISSR markers may be attributed to the differences in marker reproducibility and genome coverage among the two tested markers; this highlights the importance of loci number and their genome coverage in obtaining reliable assessments of genetic correlations among taxa [
21,
22,
47]; this may be helpful in genetic diversity, classification, taxonomy, systematics, and evolutionary biology [
1,
49,
50]. Our results showed that the genetic identification and classification depend mainly on the efficiency of a marker [
26,
51].
Markers had categorized by variance decomposition
C. intybus var. intybus and
C. intybus var.
foliosum into a group and
C. endivia var.
crispum and
C. endivia subsp.
pumilum into a group except those revealed by ISSR showing
C. intybus var. foliosum and
C. endivia var.
endivia into a group and
C. endivia subsp.
pumilum and
C. endivia var. crispum into a group, as shown in
Table 4. That may be due to the variance in the resolution of ISSRs and targeting different loci of the genome by these techniques [
21,
26,
51]. Earlier microsatellite markers could not differentiate between the species [
52]. Additionally, it may be due to the makeup of taxon’s genotype or the effect of evolution naturally.
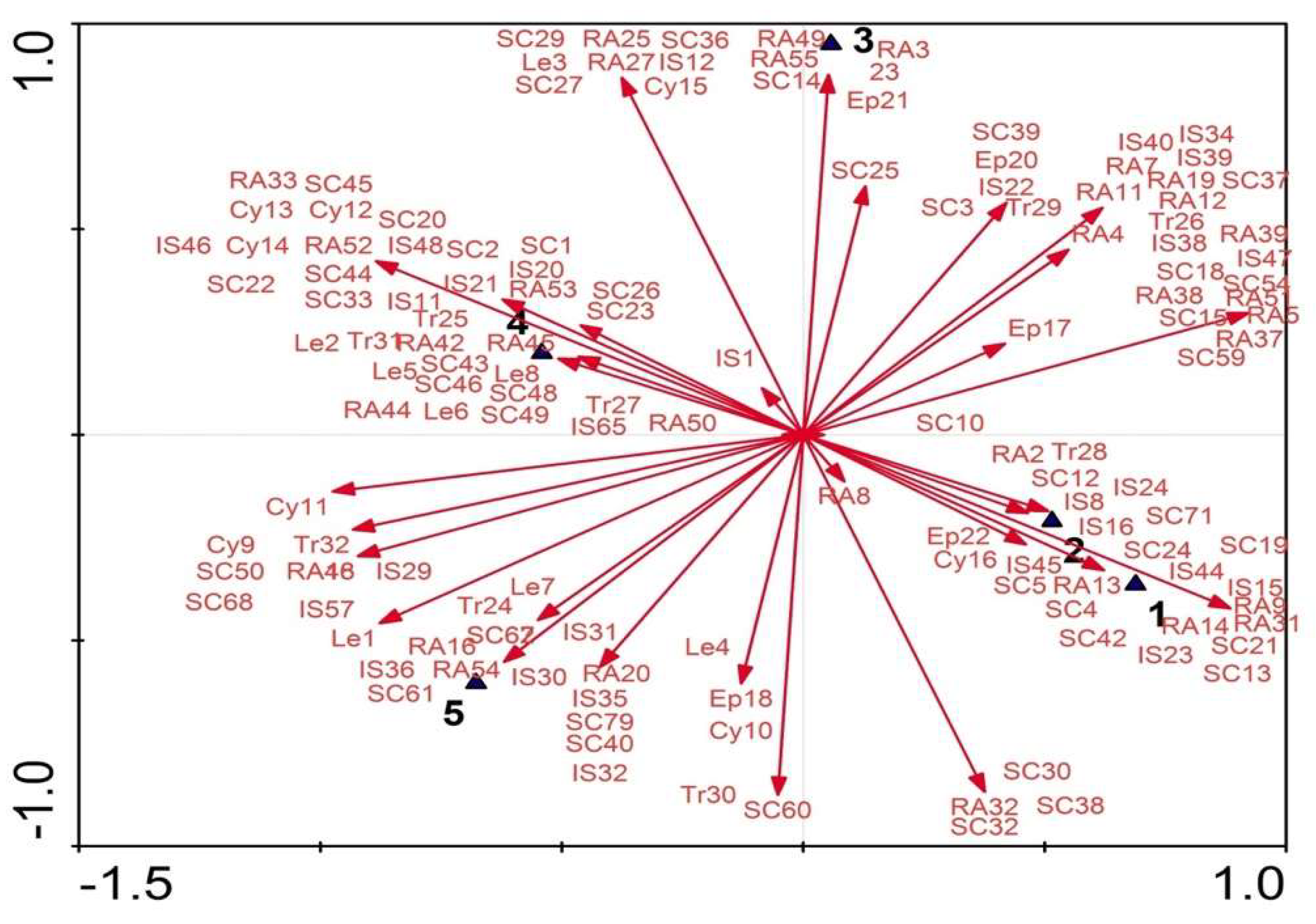
The DCCA analysis of morphological and molecular characteristics revealed that
Cichorium intybus var.
intybus is related to
C. intybus var.
foliosum, while
C. endivia var.
endivia is related to
C. endivia var.
crispum and
C. endivia subsp.
pumilum. These results are consistent with those of Kiers [
11] and Kiers et al. [
43]. In earlier publications, it has been argued that the Egyptian chicory was classified into
C. intybus [
38,
39,
40]. The integrated approaches supports a better authentication and verification of the species and taxa. According to the results of this research we argue that the Egyptian chicory is classified into
C. endivia subsp.
pumilum. 4. Materials and Methods
4.1. Plant Materials
Cypselae of Cichorium species were obtained from the collection maintained by Vegetable Production Research Department, Agricultural Research Center (ARC), Giza, Egypt. Five taxa of the genus Cichorium were used in this study. Seeds and herbarium specimens of Cichorium taxa, Cichorium intybus var. intybus, C. intybus var. foliosum, C. endivia var. endivia, C. endivia var. crispum, and C. endivia subsp. pumilum were identified under the authority of the Herbarium, Botany Dept., National Gene Bank (NGB), Agricultural Research Center (ARC), Giza, Egypt.
4.2. Germination
Light intensities at mid-canopy were kept at approximately 400 µmols m2s−1 in the growth chambers for germination. A photoperiod was adjusted at 16 h of light and 8 h of darkness using fluorescent and incandescent lights. A daytime temperature of 23 °C and a nighttime temperature of 18 °C were maintained using chart recorders. The relative humidity was maintained at approximately 50%. First, the seed surface was sterilized using a solution of 5% sodium hypochlorite for 10 min and rinsed several times with sterile distilled water. Next, 10 seeds of each taxon were replicated in fours on a blotter, to which 10 mL of the test solution was added. Seeds germinated in the growing media containing peat moss, sandy soil, and perlite (1:1:1) under growth chamber conditions. Outer leaves at the basal nodes were taken for macromorphological and micromorphological analysis.
4.3. Microscopy
4.3.1. Scanning Electron Microscopy (SEM)
Cypselae and leaves of each
Cichorium taxon were separately cut into small sections of 4–8 mm long. Glutaraldehyde solution (6%, pH 7.3) was used to fix the sections for 12 h [
53]. These were then rinsed with a 0.05 M C
2H
6AsNaO
2 (sodium cacodylate) buffer (pH 7.5) and rinsed with distilled water. This was then dehydrated with gradual ethanol concentrations (10–100%) for 20 min for each concentration. A Hitachi HCP-2 critical point dryer was used to dry the samples, which were then mounted onto aluminum stubs with carbon-coated sputtering (Elko IB-3 Ion Coater). JEOL (JSM-6390 LV) SEM, Model JEOLJSM- 5500 LV was used to examine the samples. As in SEM, both fixation and dehydration procedures were conducted using energy-dispersive X-ray spectroscopy (EDX). NORAN System SIX software (V.1.8) was used for photomicrographs digitally taken at the Electron Microscopy Unit at the National Research Centre, Dokki, Egypt. The terminology for the cypsela, abaxial epidermis, and stomata used in this study follows Barthlott [
54], Garg and Sharma [
55], Mukherjee and Nordenstam [
56], and Dilcher [
57]. In addition, all morphological characters regarding the leaf, cypsela, and trichomes were documented to create classical keys.
4.3.2. Light Microscopy
A piece of the middle part of the leaf (1 cm
2) from each plant was taken and dehydrated in a series of ethanol concentrations (50–100%). The specimens were then embedded in paraffin wax (m.p. 58–61 °C) by xylol (solvent), sectioned at 15 μm on a Jung PM 2045 rotary microtome, and mounted on glass slides with egg albumen (adhesive agent). The wax was dissolved in xylol, and the slides were stained using light green and safranin. Using Canada balsam (mounting agent), permanent slides were prepared [
58,
59]. A digital camera was used to capture the photomicrographs.
4.4. Morphology
Twenty eight traits are presented including thirteen attributes of leaves and fifteen attributes of cypsela; the traits were estimated from approximately ten healthy plants from each taxon and are presented according to the relevant terminology [
60].
4.5. DNA Extraction
The gDNA extraction was performed by the manufacturer’s instructions of Zymo extraction kit (Zymo Research, Inc., Irvine, CA, USA). The DNA purity and quantity were checked via nanodrop and then stored for PCR analysis. The integrity of gDNA was verified by agarose gel electrophoresis in a 1% agarose/1 × TAE gel containing 1 × Sybr® Safe DNA gel stain (Life Technology, Carlsbad, CA, USA). The isolated genomic DNA samples were diluted to 10 ng/μL. Then, good-quality gDNA samples were used for PCR amplification.
4.6. PCR-Based Markers
Three PCR-based markers were used for providing more information on
Cichorium’s genetic relatedness which are random amplified polymorphic DNA (RAPD), Inter Simple Sequence Repeats (ISSR), and start codon target (SCoT), as given in
Table 3. RAPD-PCR amplification was carried out with ten synthesized random 10-mer arbitrary primers (Operon Biotechnologies, Inc., Ebersberg, Germany). The procedure was done as detailed in Williams et al. [
20] and Williams and St Clair [
30] with minor modifications. The amplification by ISSR to detect polymorphisms among accession for both species was carried out as described by Yang and Park [
61] with slight modifications. The procedure of SCoT was designed by describing of Collard and Mackill [
62], which synthesized by Operon Biotechnologies, Inc., GmbH, (Cologne, Germany).
The PCR reactions were performed in a 25 μL reaction mixture that contained 25 ng of template DNA, 0.2 μM dNTPs, 1 µM of each tested primer, 1.5 mM MgCl
2, 1× PCR buffer, and 1 U of Go-Taq Flexi polymerase. In the initial denaturation cycle, the PCR program was adjusted at 94 °C for 5 min, then followed by 35 cycles. Each cycle involved 94 °C for 1 min, varying annealing temperature for each primer (
Table 3) for 1 min, then 72 °C for 90 s, and 72 °C for 7 min in the final step for extension. The PCR amplification reaction products (Amplicons) were resolved in 1.0–1.5% agarose gel that contained ethidium bromide (0.5 µg/mL) in 1× TBE as a running buffer. A 100 bp plus DNA Ladder was used as molecular size standards.
PCR products were run on agarose gel in 1% agarose × TAE gel containing 1 × Sybr® Safe DNA gel stain (Life Technology, Carlsbad, CA, USA) at 100 V for 30 min. Gels were visualized and photographed with a Gel DocTM XR+ System with Image LabTM Software (Bio-Rad®). Amplicon banding profiles were scored as present (1) or absent (0) in a binary matrix based on standard markers using Alpha Ease FCTM (version 4.0.1) software.
4.7. Statistical Analysis
Similarity for a binary matrix was estimated according to the Jaccard coefficient [
63]. The dendrogram generated the Un-weighted Pair Group Method algorithms with Arithmetic (UPGMA) averages according to Nei and Li [
64] to determine the genetic relationship among taxa. In addition, a Detrended Canonical Correspondence Analysis (DCCA) was performed using CANOCO V. 4.5 and CanoDraw V. 4.1. The dataset was entered into SPSS (version 14.0), as well as the add-in packages of StatistiXL (Kovach Computing Service 2013, version 1.8:
http://www.xlstat.com (last accessed on 7 February 2022) and GenAlEx Genetic Analysis (version 6.5) in Microsoft Excel [
65,
66].
5. Conclusions
The integration of various identification methods, including macro- and micro-morphological and molecular characterization, plays a chief role in the identification and authentication of plant genera, species, and taxa. Although measuring the variability of morphological attributes does not provide extensive information, it can still be helpful for breeding and improvement programs. This is especially important in changing climates, including the Mediterranean region. The existence of these distinct taxa and their phylogenetic relationships were assessed and confirmed in this study using the phenological, and molecular markers, and phylogenetic methodologies. The approaches used in this study proved that these tools are helpful for phylogenetic studies at the species level or higher taxonomic ranks in the genus Cichorium.
We report that the leaf shape and type, the cypsela anticlinal and periclinal walls, pappus bristles, the epidermal cell wall, and the stomatal type were the most critical characteristics in the construction of the dichotomous indented key for Cichorium taxa. PCR-based identification confirmed these results revealed through the phylogenic tree. We observe that the integration of phenological and genetic characterization is helpful in the authentication of Cichorium taxa. Further research could focus on intra-species variability, diversity, and genetic differences among individuals of the Cichorium taxa. Despite the success of the molecular marker approach, we recommend the use of supplemental techniques such as DNA barcoding for improved classification and identification, along with more core and base collections of the studied plant materials. Our observations suggest that these taxa have the potential to provide rich genetic resources for further research in genetic biodiversity, conservation, and plant breeding programs. The chicory genetic assembly among different countries in the Mediterranean region visibly diverges with a firm amount of gene flow. The PCR-based markers could be utilized for analyzing genetic relationships and taxonomies of Cichorium taxa. Finally, it could be concluded that the Egyptian chicory belongs to the C. endivia subsp. pumilum.

 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}