
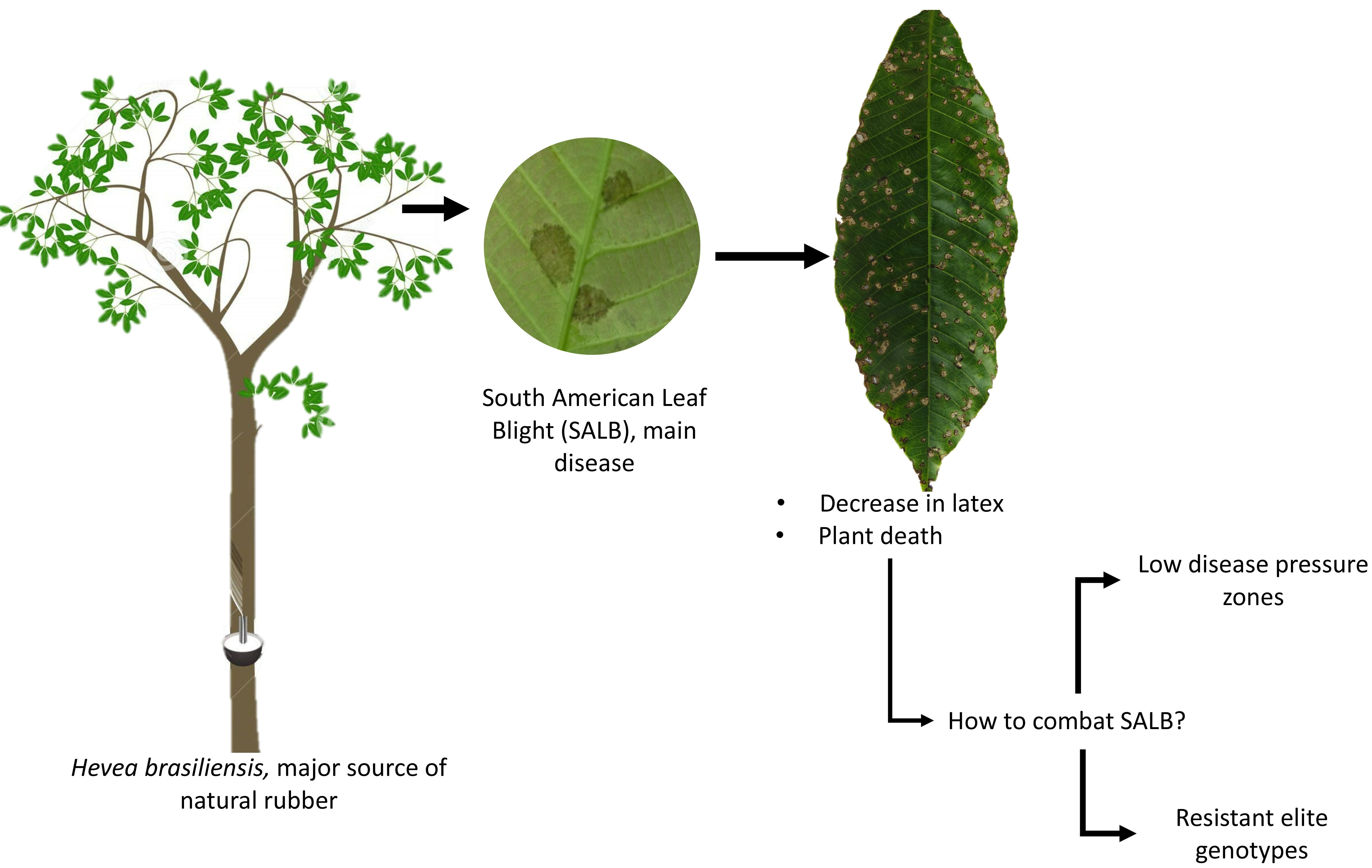
Performance- and Resistance-Related Early Responses of Colombian Elite Rubber Tree Genotypes under Low Pressure of South American Leaf Blight: Implications for Disease Management in the Amazon
 , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Climate
2.2. Effects of the Fixed Factors
2.3. Growth and Early Yield
2.4. Resistance to SALB
3. Discussion
4. Materials and Methods
4.1. Study Area
4.1.1. Location
4.1.2. Soils
4.1.3. Climate
4.2. Experimental Design and Plot Management
4.3. Plant Material
4.4. Evaluations and Data Analysis
4.4.1. Growth and Early Yield
4.4.2. Resistance to SALB
4.4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rivano, F.; Mattos, C.R.R.; Cardoso, S.E.A.; Martinez, M.; Cevallos, V.; Le Guen, V.; Garcia, D. Breeding Hevea brasiliensis for Yield, Growth and SALB resistance for high disease environments. Ind. Crops Prod. 2013, 44, 659–670. [Google Scholar] [CrossRef]
- Mazlan, S.; Jaafar, N.; Wahab, A.; Sulaiman, Z.; Rajandas, H.; Zulperi, D. Major diseases of rubber (Hevea brasiliensis) in Malaysia. Pertanika J. Sch. Res. Rev. 2019, 5, 10–21. [Google Scholar]
- Prabhakaran, K.P. The Agronomy and Economy of Important Tree Crops of the Developing World. Nair, K.P.P., Ed.; Elsevier: New Delhi, India, 2010; ISBN 978-0-12-384677-8. [Google Scholar]
- Food and Agriculture Organization—FAO FAOSTAT. 2023. Available online: http://www.fao.org/faostat/ (accessed on 13 September 2023).
- de Castro Sant’Anna, I.; Gouvêa, L.R.L.; Martins, M.A.; Scaloppi, E.J., Jr.; de Freitas, R.S.; Gonçalves, P. Genetic diversity associated with natural rubber quality in elite genotypes of the rubber tree. Sci. Rep. 2021, 11, 1081. [Google Scholar] [CrossRef]
- Van Truong, V.; Van Lam, L.; Thuy, L.M.; Dinh, H.D.; Thao, N.T.; Rivallan, R.; Van Biet, H.; Guen, V. Le population genetic structure of a thousand rubber tree accessions from wild Rondônia populations conserved in Vietnam. Genet. Resour. Crop Evol. 2020, 67, 475–487. [Google Scholar]
- Maiden, N.A.; Syd Ali, N.; Ahmad, K.; Atan, S.; Wong, M.Y. Growth and physiological responses of Hevea brasiliensis to Rigidoporus microporus infection. J. Rubber Res. 2022, 25, 213–221. [Google Scholar] [CrossRef]
- Malmonge, J.A.; Camillo, E.C.; Moreno, R.M.B.; Mattoso, L.H.C.; McMahan, C.M. Comparative study on the technological properties of latex and natural rubber from Hancornia speciosa Gomes and Hevea brasiliensis. J. Appl. Polym. Sci. 2008, 111, 2986–2991. [Google Scholar] [CrossRef]
- Jaimes, Y.; Rojas, J.; Cilas, C.; Furtado, E.L. Suitable climate for rubber trees affected by the South American Leaf Blight (SALB): Example for identification of escape zones in the Colombian Middle Magdalena. Crop Prot. 2016, 81, 99–114. [Google Scholar] [CrossRef]
- Tapiero, A.L. Enfermedades y su control. In Modelo Productivo Para el Cultivo de Caucho Natural (Hevea brasiliensis) en el Bajo Cauca Antioqueño y en el sur de Córdoba; Gutiérrez Vanegas, A.J., Correa Pinilla, D.E., Gómez Valderrama, J.A., Amado Saavedra, G.M., Martínez Atencia, J., Álvarez-Láinez, M.L., Córdoba Gaona, O.d.J., Tapiero Ortiz, A.L., Monsalve García, D.A., Gil Restrepo, J.P., Eds.; AGROSAVIA: Mosquera, Colombia, 2022; pp. 145–162. [Google Scholar]
- de Araújo, A.E.; Filho, A.Ν.; Nóbrega, M.Β.M.; Sousa, N.R.; Santos, J.W. dos Reação de Dez Clones de Seringueira (Hevea benthamiana) a três isolados de Microcyclus ulei. Acta Amaz. 2001, 31, 349. [Google Scholar] [CrossRef]
- Junqueira, N.T.V.; Gasparotto, L.; Lieberei, R.; Normando, M.C.S.; Lima, M.I.P.M. Especialização fisiológica de Microcyclus ulei em diferentes espécies de Seringueira: Identificação de grupos de patótipos. Fitopatol. Bras. 1989, 14, 205. [Google Scholar]
- Gasparotto, L.; Lieberei, R.; Trindade, D. In vitro conidia germination of Microcyclus ulei and its sensitivity to fungicides. Fitopatol. Bras. 1984, 9, 505–511. [Google Scholar]
- Gasparotto, L.; Pereira, J.C.R.; Furtado, E.L. Capítulo 3: Doenças das folhas. In Doenças da Seringueira no Brasil; Embrapa: Brasilia, Brazil, 2012; pp. 39–176. [Google Scholar]
- Sterling, A.; Rodríguez, O.L.; Correa, J. Evaluación fitosanitaria y resistencia en campo a Microcyclus ulei de Hevea Brasiliensis en la Amazonia Colombiana. Rev. Colomb. Amaz. 2009, 5, 80–90. [Google Scholar]
- Garzón, C. Principales Enfermedades y Plagas en el Cultivo de Caucho (Hevea brasiliensis) con Énfasis en la Amazonia Colombiana; Instituto Amazónico de Investigaciones Científicas SINCHI: Bogotá, Colombia, 2000. [Google Scholar]
- Jaimes, Y.Y.; Rojas, J.; Furtado, E.L. Clones de Hevea brasiliensis de alta productividad caracterizados por resistencia a Microcyclus ulei en jardin clonal en el Magdalena Medio Colombiano. Summa Phytopathol. 2015, 41, 115–120. [Google Scholar]
- Chee, K.H.; Holliday, P. South American Leaf Blight of Hevea rubber. Malaysian Rubber Res. Dev. Board Monogr. 1986, 13, 50. [Google Scholar]
- Lieberei, R. South American Leaf Blight of the Rubber Tree (Hevea spp.): New steps in plant domestication using physiological features and molecular markers. Ann. Bot. 2007, 100, 1125–1142. [Google Scholar] [CrossRef]
- da Hora Júnior, B.T.; de Macedo, D.M.; Barreto, R.W.; Evans, H.C.; Mattos, C.R.R.; Maffia, L.A.; Mizubuti, E.S.G. Erasing the Past: A New Identity for the samoclean pathogen causing South American Leaf Blight of rubber. PLoS ONE 2014, 9, e104750. [Google Scholar] [CrossRef]
- Le Guen, V.; Garcia, D.; Mattos, C.R.R.; Doaré, F.; Lespinasse, D.; Seguin, M. Bypassing of a polygenic Microcyclus ulei resistance in rubber tree, analyzed by QTL detection. New Phytol. 2007, 173, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Sobha, S.; Rekha, K.; Uthup, T.K. Abstract Biotechnological Advances in rubber tree (Hevea brasiliensis Muell. Arg.) breeding. In Advances in Plant Breeding Strategies: Industrial and Food Crops; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer Nature: Cham, Switzerland, 2019; Volume 6, pp. 179–236. [Google Scholar]
- Correa-Pinilla, D.E.; Gutiérrez-Vanegas, A.J.; Gil-Restrepo, J.P.; Martínez-Atencia, J.; Córdoba-Gaona, O.d.J. Agroecological and South American Leaf Blight Escape Zones for rubber cultivation in Colombia. Agron. J. 2022, 114, 2830–2844. [Google Scholar] [CrossRef]
- Rivano, F.; Vera, J.; Cevallos, V.; Almeida, D.; Maldonado, L.; Flori, A. Performance of 10 Hevea brasiliensis Clones in Ecuador, under South American Leaf Blight escape conditions. Ind. Crops Prod. 2016, 94, 762–773. [Google Scholar] [CrossRef]
- Rivano, F.; Maldonado, L.; Simbaña, B.; Lucero, R.; Gohet, E.; Cevallos, V.; Yugcha, T. Suitable Rubber Growing in Ecuador: An approach to South American Leaf Blight. Ind. Crops Prod. 2015, 66, 262–270. [Google Scholar] [CrossRef]
- Castañeda, A. Zonas Aptas Para el Cultivo de Caucho en Colombia; Serie Técnica No. 39; CONIF: Bogotá, Colombia, 1997. [Google Scholar]
- Pilau, F.G.; Marin, F.R.; de Camargo, M.B.P.; Assad, E.D.; Pinto, H.S.; Barbarisi, B.F. Zoneamento agroclimático da heveicultura para as regiões Sudeste e Centro-Oeste Do Brasil. Rev. Bras. Agrometeorol. Piracicaba 2010, 15, 161–168. [Google Scholar]
- Carmo, C.A.F.d.S.d.; Lumbreras, J.F.; Naime, U.J.; Gonçalves, A.O.; Fidalgo, E.C.C.; Ágilo, M.L.D.; Lima, J.A.d.S. Aspectos culrurais e zoneamento da Seringueirano Estado Do Rio de Janeiro. Bol. Pesqui. Desenvolv. 2004, 60, 49. [Google Scholar]
- Sterling, A.; Pimentel-Parra, G.A.; Virguez-Díaz, Y.R.; Suárez-Córdoba, Y.D.; Hoyos-Duarte, J.D.; Fonseca-Restrepo, J.A. Long-Term resistance in promising rubber tree genotypes as a breeding source for improving South American Leaf Blight management under high disease incidence in the Colombian Amazon. Crop Prot. 2021, 150, 105817. [Google Scholar] [CrossRef]
- Sterling, A.; Galindo-Rodríguez, L.C.; Suárez-Córdoba, Y.D.; Velasco-Anacona, G.; Andrade-Ramírez, T.; Gómez-Torres, A.K. Early assessing performance and tesistance of Colombian Rubber Tree Genotypes under High South American Leaf Blight Pressure in Amazon. Ind. Crops Prod. 2019, 141, 111775. [Google Scholar] [CrossRef]
- Sterling, A.; Martínez-Viuche, E.J.; Suárez-Córdoba, Y.D.; Agudelo-Sánchez, A.A.; Fonseca-Restrepo, J.A.; Andrade-Ramírez, T.K.; Virguez-Díaz, Y.R. Assessing Growth, early yielding and resistance in rubber tree clones under low South American Leaf Blight pressure in the Amazon Region, Colombia. Ind. Crops Prod. 2020, 158, 112958. [Google Scholar] [CrossRef]
- Sterling, A.; Rodríguez, C.H. Valoración de Nuevos Clones de Hevea brasiliensis con Proyección Para la Amazonia Colombiana: Fases de pre y Post-Sangría Temprana en el Caquetá; Sterling, A., Rodríguez, C.H., Eds.; Instituto Amazónico de Investigaciones Científicas—SINCHI: Bogotá, Colombia, 2020. [Google Scholar]
- Sterling, A.; Rodríguez, C.H. Estrategias de Manejo Para las Principales Enfermedades y Plagas del Cultivo del Caucho con Énfasis en La Amazonia Colombiana; Instituto Amazónico de Investigaciones Científicas—SINCHI: Florencia Caquetá, Colombia, 2018. [Google Scholar]
- Sterling, A.; Martínez-Viuche, E.J.; Pimentel-Parra, G.A.; Suárez-Córdoba, Y.D.; Fonseca-Restrepo, J.A.; Virguez-Díaz, Y.R. Dynamics of adaptive responses in growth and resistance of rubber tree clones under South American Leaf Blight non-escape conditions in the Colombian Amazon. Ind. Crops Prod. 2019, 141, 111811. [Google Scholar] [CrossRef]
- Verbeke, G.; Molenberghs, G. Linear Mixed Models for Longitudinal Data; Springer: New York, NY, USA, 2000. [Google Scholar]
- Rivano, F.; Martinez, M.; Cevallos, V.; Cilas, C. Assessing resistance of rubber tree clones to Microcyclus ulei in large-scale clone trials in Ecuador: A less time-consuming field method. Eur. J. Plant Pathol. 2010, 126, 541–552. [Google Scholar] [CrossRef]
- Meenakumari, T.; Meenattoor, J.R.; Thirunavoukkarasu, M.; Vinod, K.K.; Krishan, B.; Gireesh, T.; Thomas, V.; Mydin, K.K.; Jacob, J. Dynamics of long-term adaptive responses in growth and rubber yield among Hevea brasiliensis genotypes introduced to a dry sub-humid climate of eastern India. Ind. Crops Prod. 2018, 119, 294–303. [Google Scholar] [CrossRef]
- Da Silva, K.R.; Cecílio, R.A.; Xavier, A.C.; Pezzopane, J.R.M.; de Oliveira Garcia, E. Zoneamento edafoclimático para a cultura da Seringueira No Espírito Santo. IRRIGA 2013, 18, 1–12. [Google Scholar] [CrossRef]
- Guyot, J.; Condina, V.; Doaré, F.; Cilas, C.; Sache, I. Segmentation Applied to Weather-Disease Relationships in South American Leaf Blight of the Rubber Tree. Eur. J. Plant Pathol. 2010, 126, 349–362. [Google Scholar] [CrossRef]
- Sterling, A.; Suárez, J.C.; Rodríguez, C.H.; Caicedo, D.F.; Salas, Y.M.; Virguez, Y.R. Crecimiento inicial de clones promisorios de Hevea brasiliensis (Willd. Ex A. Juss.) Müll. Arg. en sistema agroforestal en Caquetá, Colombia. Colomb. For. 2015, 18, 175. [Google Scholar] [CrossRef]
- Gireesh, T.; Meenakumari, T.; Mydin, K.K. Fast Track Evaluation and selection of Hevea brasiliensis clones from a clonal nursery. Ind. Crops Prod. 2017, 103, 195–201. [Google Scholar] [CrossRef]
- Gonçalves, P.D.S.; Bortoletto, N.; Furtado, E.L. Desempenho de Clones de Seringueira da Série IAC 300 Selecionados Para a Região Noroeste Do Estado de São Paulo; Embrapa Secretaria de Pesquisa e Desenvolvimento, Pesquisa Agropecuária Brasileira: Brasília, Brazil, 2001; pp. 589–599. [Google Scholar]
- Vrignon-Brenas, S.; Gay, F.; Ricard, S.; Snoeck, D.; Perron, T.; Mareschal, L.; Laclau, J.P.; Gohet, É.; Malagoli, P. Nutrient management of immature rubber plantations. A review. Agron. Sustain. Dev. 2019, 39, 11. [Google Scholar] [CrossRef]
- Gouvêa, L.R.L.; Silva, G.A.P.; Verardi, C.K.; Oliveira, A.L.B.; Gonçalves, P. Simultaneous selection of rubber yield and girth growth in young rubber trees. Ind. Crops Prod. 2013, 50, 39–43. [Google Scholar] [CrossRef]
- Gonçalves, P.d.S.; Silva, M.d.A.; Gouvêa, L.R.L.; Scaloppi, E.J., Jr. Genetic variability for girth growth and rubber yield in Hevea brasiliensis. Sci. Agric. 2006, 63, 246–254. [Google Scholar] [CrossRef]
- Vélez-Sánchez, D.F.; Nieto-Rodríguez, V.M. Desempeño inicial de clones de caucho en el municipio de Puerto Carreño, Vichada, Colombia. Colomb. For. 2013, 16, 186–199. [Google Scholar]
- Vinod, K.K.; Rajeswari, M.J.; Pothen, J.; Krishnakumar, A.K.; Sethuraj, M.R. Performance Analysis for Wintering Pattern in Hevea brasiliensis Clones. Ind. J. Nat. Rubber Res. 1996, 9, 44–47. [Google Scholar]
- Quesada-Méndez, I.; Aristizábal-Gutiérrez, F. Characterization of two latex parameters from Hevea Brasiliensis (Willd. Ex A. Juss.) Müll. Arg. Clones in the Colombian Altillanura. Colomb. For. 2012, 15, 139–146. [Google Scholar] [CrossRef]
- Sethuraj, M.R. Yield Components in Hevea brasiliensis. In Natural Rubber Biology, Cultivation and Technology; Sethuraj, M.R., Mathew, N.M., Eds.; Developments in Crop Science; Elsevier: Kerala, India, 1992; Volume 23, pp. 137–163. [Google Scholar]
- Mendoza-Vargas, L.P.; Jiménez-Forero, J.A. Effect of inclusion of anticoagulants during the benefit of Hevea brasiliensis (Willd. Ex a.Juss.) Müll.Arg. Colomb. For. 2018, 21, 91–101. [Google Scholar] [CrossRef]
- Ajith, S.V.; Nair, N.N.; Sathik, M.B.M.; Meenakumari, T. Breeding for low-temperature stress tolerance in Hevea brasiliensis: Screening of newly developed clones using latex biochemical parameters. J. Rubber Res. 2021, 24, 819–828. [Google Scholar] [CrossRef]
- Cáceres, S.A.P.; Gauthier-Maradei, P. Thermogravimetric analysis as a new method to determine of total solid content (TSC) and dry rubber content (DRC) of natural latex. Rev. ION 2012, 25, 57–65. [Google Scholar]
- Támara, A.M.; Reyes, H. Physicochemical analysis and characterization of the Hevea brasiliensis species. Entre Cienc. Ingienería 2015, 18, 9–15. [Google Scholar]
- Jacob, J.; Prévöt, J.C.; Roussel, D.; Lacrotte, R.; Serres, E.; d’Auzac, J.; Eschbach, J.M.; Omont, H. Yield limiting factors for latex yield of Hevea. In Physiology of Rubber Tree Latex. The Laticiferous Cell and Latex: A Model of Cytoplasm; D’Auzac, J., Jacob, J.-L., Chrestin, H., Eds.; CRC Press: Boca Raton, FL, USA, 1989; Volume 1, pp. 345–397. [Google Scholar]
- Guyot, J.; Condina, V.; Doaré, F.; Cilas, C.; Sache, I. Role of Ascospores and Conidia in the initiation and spread of South American Leaf Blight in a rubber tree plantation. Plant Pathol. 2014, 63, 510–518. [Google Scholar] [CrossRef]
- Guyot, J.; Le Guen, V. A Review of a century of studies on South American Leaf Blight of the rubber tree. Plant Dis. 2018, 102, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Le Guen, V.; Garcia, D.; Mattos, C.; Fouet, O.; Doaré, F.; Condina, V.; Seguin, M. A newly identified locus controls complete resistance to Microcyclus ulei in the Fx2784 rubber clone. Tree Genet. Genomes 2013, 9, 805–812. [Google Scholar] [CrossRef]
- Instituto Geográfico Agustín Codazzi (IGAC) Estudio General de Suelos y Zonificación de Tierras Departamento de Caquetá Escala 1.100.000; Imprenta Nacional de Colombia: Bogotá, Colombia, 2014.
- Instituto Geográfico Agustín Codazzi-IGAC Caquetá. Características Geográficas; IGAC: Bogotá, Colombia, 2010. [Google Scholar]
- Clément-Demange, A.; Nicolas, D.; Legnaté, H.; Rivano, F.; Le Guen, V.; Gnagne, M.; Chapuset, T. Hévéa: Stratégies de sélection. Plant. Rech. Dev. 1995, 2, 5–14. [Google Scholar]
- Sterling, A.; Andrade-Ramírez, T.K.; Clavijo-Arias, E.A.; Guaca-Cruz, L.; Celis-Hernández, M.D.; Noriega, J.E.; Torres-Barrero, R.; Suárez-Salazar, J.C. evaluación de los rasgos de crecimiento, nutricionales y fisiológicos de genotipos caquetenses selección ECC 1 de caucho (Hevea brasiliensis) en campos clonales a gran escala en el Caquetá. In Valoración de Nuevos Clones de Hevea brasiliensis con Proyección Para la Amazonia Colombiana: Fases de pre y Post-Sangría Temprana en el Caquetá; Sterling, A., Rodríguez, C.H., Eds.; Instituto Amazónico de Investigaciones Científicas SINCHI: Bogotá, Colombia, 2020; pp. 127–152. [Google Scholar]
- Sterling, A.; Rodríguez, C.H. Nuevos Clones de Cuacho Natural Para la Amazonía Colombiana: Énfasis en la Resistencia al mal Suramericano de las Hojas (Mycrocyclus ilei), 1st ed.; Sterling, A., Rodríguez, C.H., Eds.; Instituto Amazónico de Investigaciones Científicas SINCHI: Bogotá, Colombia, 2011. [Google Scholar]
- Confederación Cauchera Colombiana (CCC). Estado Actual Del Gremio Cauchero Colombiano; CAELCA: Bogotá, Colombia, 2015. [Google Scholar]
- Silva, G.A.P.; Gouvêa, L.R.L.; Verardi, C.K.; de Oliveira, A.L.B.; Gonçalves, P.d.S. Annual growth increment and stability of rubber yield in the tapping phase in rubber tree clones: Implications for early selection. Ind. Crops Prod. 2014, 52, 801–808. [Google Scholar] [CrossRef]
- Lacote, R.; Gabla, O.; Obouayeba, S.; Eschbach, J.M.; Rivano, F.; Dian, K.; Gohet, E. Long-Term Effect of ethylene stimulation on the yield of rubber trees is linked to latex cell biochemistry. Field Crop Res. 2010, 115, 94–98. [Google Scholar] [CrossRef]
- Junqueira, N.T.V.; Chaves, G.M.; Zambolim, L.; Gasparotto, L.; Alfenas, A.C. Variabilidade fisiológica de Microcyclus ulei. Fitopatol. Bras. 1986, 11, 823–833. [Google Scholar]
- Mattos, C.; Garcia, D.; Pinard, F.; Le Guen, V. Variabilidade de isolados de Microcyclus ulei No Sudeste de Bahia. Fitopatol. Bras. 2003, 5, 502–507. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; Saikat, D.; Sarkar, D.; R Development Core Team. Nlme: Linear and Nonlinear Mixed Effects Models; R Core Team: Vienna, Austria, 2013; Volume 3, pp. 1–109. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarine, M.G.; Gonzales, L.; Tablada, M.; Robledo, C.W. InfoStat, versión 2018; National University of Cordoba: Córdoba, Argentina, 2018.
- Doledec, S.; Chessel, D. Co-inertia Analysis: An alternative method for studying species–environment relationships. Freshw. Biol. 1994, 31, 277–294. [Google Scholar] [CrossRef]
- Thioulouse, J.; Chessel, D.; Dolédec, S.; Olivier, J.-M. ADE-4 a Multivariate analysis and graphical display software. Stat. Comput. 1997, 7, 75–83. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Compound Symmetric Model with Heterogeneous Variances (CSH) | First-Order Autoregressive Model with Heterogeneous Variances (ARH1) | First-Order Continuous Autoregressive Model with Heterogeneous Variances (CARH1) | Unstructured Model with Heterogeneous Variances (UNH) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | Log Lik | AIC | BIC | Log Lik | AIC | BIC | Log Lik | AIC | BIC | Log Lik | AIC | BIC |
| CTpt | −175.38 | 420.77 | 508.26 | −170.39 | 410.77 | 498.27 | −170.39 | 410.77 | 498.27 | −165.18 | 404.35 | 496.84 |
| AT1 | 159.78 | −235.6 | −130.6 | 159.97 | −236 | −131 | 159.97 | −235.95 | −131 | 161.4 | −234.8 | −124.81 |
| AT2 | 63.18 | −56.36 | 31.13 | 63.29 | −56.59 | 30.91 | 63.29 | −56.59 | 30.91 | 64.22 | −54.44 | 38.06 |
| ST | 48.8 | −27.59 | 59.9 | 48.68 | −27.37 | 60.13 | 48.68 | −27.36 | 60.13 | 49.65 | −25.3 | 67.2 |
| TR | 97.9 | −125.8 | −38.31 | 97.1 | −124.2 | −36.7 | 97.1 | −124.19 | −36.7 | 101.28 | −128.57 | −36.07 |
| Variable | DF1 | DF2 | DF3 | Genotype | Year | Genotype × Year |
|---|---|---|---|---|---|---|
| CTpt | 9 | 2 | 18 | 4.42 (0.0001) | 714.72 (<0.0001) | 2.23 (0.0074) |
| AT1 | 9 | 2 | 18 | 5.35 (<0.0001) | 89.39 (<0.0001) | 2.98 (0.0004) |
| AT2 | 9 | 2 | 18 | 16.19 (<0.0001) | 256.90 (<0.0001) | 2.68 (0.0012) |
| ST | 9 | 2 | 18 | 19.30 (<0.0001) | 215.16 (<0.0001) | 3.08 (0.0002) |
| TR | 9 | 2 | 18 | 3.50 (0.0010) | 116.21 (<0.0001) | 2.09 (0.0125) |
| DRC | 9 | - | - | 10.14 (<0.0001) | - | - |
| DRY | 9 | - | - | 18.85 (<0.0001) | - | - |
| CTo | 9 | - | - | 5.69 (<0.0001) | - | - |
| Genotype | Year 1 | Year 2 | Year 3 |
|---|---|---|---|
| ECC 25 | 9.37 ± 0.49 a,b k | 18.33 ± 0.88 def | 25.75 ± 1.24 ab |
| ECC 29 | 9.43 ± 0.49 k | 16.98 ± 0.88 fgh | 22.80 ± 1.24 bc |
| ECC 35 | 7.78 ± 0.49 l | 13.33 ± 0.88 i | 20.23 ± 1.24 cde |
| ECC 60 | 9.03 ± 0.49 k | 14.89 ± 0.88 hi | 21.44 ± 1.24 c |
| ECC 64 | 10.69 ± 0.49 j | 20.44 ± 0.88 cd | 27.48 ± 1. 24 a |
| ECC 66 | 8.73 ± 0.49 kl | 15.31 ± 0.88 ghi | 22.54 ± 1.24 bc |
| ECC 73 | 9.44 ± 0.49 k | 16.66 ± 0.88 fgh | 22.84 ± 1.24 bc |
| ECC 83 | 9.62 ± 0.49 jk | 17.31 ± 0.88 efg | 23.61 ± 1.24 bc |
| ECC 90 | 9.02 ± 0.49 k | 15.83 ± 0.88 gh | 23.63 ± 1.24 bc |
| IAN 873 | 9.82 ± 0.49 jk | 16.28 ± 0.88 fgh | 22.41 ± 1.24 bc |
| Genotype | CTo (cm) | DRY (g.tree−1.tap−1) | DRC (%) |
|---|---|---|---|
| ECC 25 | 26.86 ± 0.35 a,b b | 5.57 ± 0.43 a | 32.45 ± 0.44 a |
| ECC 29 | 26.49 ± 0.37 b | 0.74 ± 0.18 c | 31.44 ± 2.29 a |
| ECC 35 | 25.00 ± 2.45 b | 0.42 ± 1.14 c | 26.24 ± 0.76 b |
| ECC 60 | 25.03 ± 0.93 b | 1.21 ± 0.43 c | 30.43 ± 0.70 ab |
| ECC 64 | 27.92 ± 0.26 a | 3.76 ± 0.66 b | 31.38 ± 1.01 a |
| ECC 66 | 25.54 ± 0.78 b | 1.09 ± 0.43 c | 30.75 ± 1.10 a |
| ECC 73 | 25.62 ± 0.56 b | 2.23 ± 0.43 bc | 32.42 ± 1.18 a |
| ECC 83 | 26.09 ± 0.45 b | 3.30 ± 0.24 b | 30.53 ± 1.98 ab |
| ECC 90 | 26.94 ± 0.57 b | 2.96 ± 0.81 bc | 28.83 ± 3.05 ab |
| IAN 873 | 25.47 ± 0.60 b | 1.88 ± 0.28 c | 32.83 ± 0.10 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galindo-Rodríguez, L.C.; Sterling, A.; Muñoz-Ramirez, H.; Fonseca-Restrepo, J.A. Performance- and Resistance-Related Early Responses of Colombian Elite Rubber Tree Genotypes under Low Pressure of South American Leaf Blight: Implications for Disease Management in the Amazon. Plants 2023, 12, 3627. https://doi.org/10.3390/plants12203627
Galindo-Rodríguez LC, Sterling A, Muñoz-Ramirez H, Fonseca-Restrepo JA. Performance- and Resistance-Related Early Responses of Colombian Elite Rubber Tree Genotypes under Low Pressure of South American Leaf Blight: Implications for Disease Management in the Amazon. Plants. 2023; 12(20):3627. https://doi.org/10.3390/plants12203627
Chicago/Turabian StyleGalindo-Rodríguez, Lyda Constanza, Armando Sterling, Herminton Muñoz-Ramirez, and Jesica Andrea Fonseca-Restrepo. 2023. "Performance- and Resistance-Related Early Responses of Colombian Elite Rubber Tree Genotypes under Low Pressure of South American Leaf Blight: Implications for Disease Management in the Amazon" Plants 12, no. 20: 3627. https://doi.org/10.3390/plants12203627