
The Enigmatic CA2: Exploring the Understudied Region of the Hippocampus and Its Involvement in Parkinson’s Disease

State Key Laboratory of Medical Neurobiology and MOE Frontiers Center for Brain Science, Institutes of Brain Science, Fudan University, Shanghai 200032, China
*
Author to whom correspondence should be addressed.
Biomedicines 2023, 11(7), 1996; https://doi.org/10.3390/biomedicines11071996
Submission received: 20 June 2023
/
Revised: 12 July 2023
/
Accepted: 13 July 2023
/
Published: 14 July 2023
(This article belongs to the Section Neurobiology and Clinical Neuroscience)
Abstract
:Parkinson’s disease (PD) is a neurodegenerative disease that affects both motor and non-motor functions. Although motor impairment is a prominent clinical sign of PD, additional neurological symptoms may also occur, particularly in the preclinical and prodromal stages. Among these symptoms, social cognitive impairment is common and detrimental. This article aims to review non-motor symptoms in PD patients, focusing on social cognitive deficits. It also examines the specific characteristics of the CA2 region and its involvement in social behavior, highlighting recent advances and perspectives. Additionally, this review provides critical insights into and analysis of research conducted in rodents and humans, which may help improve the understanding of the current status of putative therapeutic strategies for social cognitive dysfunction in PD and potential avenues related to the function of the hippocampal CA2 region.
1. Introduction
Parkinson’s disease (PD) is a debilitating neurodegenerative disorder characterized by the loss of dopaminergic neurons in the pars compacta of the substantia nigra, a loss that is unfortunately irreversible [1]. Studies have described many non-motor symptoms that appear in the early stages of PD [2,3], in particular, social cognitive decline such as perception, language, and decision-making [4], as well as temporal-order memory deficits [5]. Interestingly, the hippocampal CA2 region, with its unique properties, has attracted the attention of scientists due to its potential association with social cognitive functions. In this review, we first briefly describe the unique properties of the CA2 region and its putative social functions. We then review the social cognitive deficits found in PD patients and examine the evidence linking them to the CA2 region. In addition, we compare findings from rodent and human studies. Finally, we summarize potential therapeutic drugs that show promise in attenuating social cognitive dysfunction in Parkinson’s disease (PD), including those that target the CA2 region.
2. The Hippocampal Formation and the Unique Properties of the CA2 Region
The hippocampal formation (HF) is located within the medial temporal lobe in all mammalian species, close to the adjacent cerebral cortex, enabling its many crucial connections to various cortical regions. HF is a critical functional unit that contributes significantly to many vital cognitive processes in both humans and animals, including learning and memory [6], fear processing [7], spatial orientation [8], and social behavior [9]. HF includes the cornu ammonis (CA) and the dentate gyrus areas. Ramón and Cajal divided the CA into two parts: the superior region, composed predominantly of small-body neurons, and the inferior region, composed of larger vertebral-body neurons. This division was later refined by Rafael Lorente de Nó, who identified four subregions within the CA area: CA1, CA2, CA3, and CA4. He observed that the neurons in CA2 and CA3 were larger than those in CA1 and that the CA2 subregion did not receive mossy fiber projections from the DG but instead received inputs via the Schaffer collateral fibers originating in CA3 [10]. However, Dudek et al. determined that the extent to which mossy fibers project into CA2 and synapse onto CA2 pyramidal neurons is species dependent [11]. The nomenclature and function have long been subjects of controversy, with debate over whether it is a distinct or transitional region between CA1 and CA3. However, current research suggests that CA2 possesses a unique biological structure [12], which requires further investigation in humans [13]. Interestingly, researchers have also indicated that CA2 may be proportionally larger in primates than in rodents [11].
CA2 exhibits unique morphological features, including a more loosely packed stratum pyramidal in comparison to CA1. In addition, pyramidal neurons in CA2 have an oval and dense soma, which is the largest among excitatory neurons within CA regions. Neurons in CA2 are also characterized by a hyperpolarized resting membrane potential and display specific action potential firing patterns [14]. Furthermore, the afferent and efferent connections of the CA2 field have distinct origins and terminations compared to other regions of the hippocampal formation. For instance, an optogenetic study demonstrated functional monosynaptic inputs from the DG via longitudinal projections to the CA2 area [15]. Other data suggest that CA2 neurons have more extensive functional synaptic connections with the deep area of CA1 than with the superficial layer [15] and that they also exhibit stronger innervation to CA1 than to CA3 [14]. However, the degree of synaptic connectivity between layer III EC afferents and distant branches of CA2 neurons may vary between species. In particular, fibers originating from layer III EC neurons and traversing the stratum lacunosum-moleculare in the CA1 region play a role in this variation [14].
Recently, CA2 has been shown to play a central role in social behavior. Molecular markers such as Purkinje cell protein 4 (PCP4), a regulator of G protein signaling 14 protein (RGS14), and striatum-enriched protein–tyrosine phosphatase (STEP), help to identify the specific population of neurons in the CA2 region [11,12,15,16,17,18]. Lee et al. demonstrated that RGS14 deletion imparts a substantial capacity for SC-CA2 synapse, whereas wild-type CA2 neurons exhibited little LTP [16]. Researchers discovered a loss of inhibitory neurons in CA2 in a neuropsychiatric disorder-like mouse model. These mice exhibited impaired social cognition and reduced synaptic plasticity in CA2, which may be related to the loss of PV+ interneurons [19]. In addition, CA2 activates a disinhibitory circuit from the lateral septum to the ventromedial hypothalamus (LS-VMHv1), which is modulated by the signaling pathway of arginine vasopressin (AVP), a hormone and neurotransmitter, to promote social aggression [20]. Dysfunctions in this signaling pathway have also been associated with neuropsychiatric disorders such as depression, anxiety, and autism spectrum disorders. Furthermore, researchers have speculated that the CA2 region is crucial for the formation and retrieval of memories related to social encounters [21]. Although arginine vasopressin receptor 1B (AVPR1B) mRNA is highly expressed in CA2 pyramidal neurons in both humans and rodents [12,21], one study demonstrated that AVPR1B -deficient mice were unable to recognize other mice in the “social novelty test” and also showed impaired chronological-order memory [22].
Another test showed that AVPR1B knockout mice could not discriminate the object they explored and recognize its location like the control group [23]. AVPR1B deficiency in CA2 impaired social memory enhancement [24]. In addition to AVPR1B, oxytocin receptors, another social neuropeptide receptor, are also highly expressed in CA2 [25,26]. In addition, genetic evidence suggests that CA2 injury impairs social recognition in mice [22]. Interestingly, the CA2 area of the hippocampus is the only region that receives vasopressinergic input from both the paraventricular nuclei of the hypothalamus and the supramammillary nuclei (SuM)—a critical factor in the regulation of social cognitive behaviors [27,28,29,30]. Interestingly, terminals belonging to the SuM have been found to express substance P [11], which plays a central role in PD. Furthermore, research suggests that these particular SuM afferents expressing substance P specifically target CA2 in rats and have the ability to influence plasticity in pyramidal neurons located in CA2 [31]. The SuM-to-CA2 projection has also been reported in monkeys and humans and occurs during early embryonic development [32]. The reason for enhanced social performance may involve the circuit from dorsal CA2 to ventral CA1 [33], spike timing-dependent plasticity in CA2 [34], the negative regulatory role of CA2 in hippocampal sharp-wave ripples [35,36], and the distinct dendritic properties of CA2 compared to CA1 [37]. In addition, mineralocorticoid receptors (MRs) have been shown to facilitate CA2-dependent behaviors [38].
In summary, CA2 pyramidal neurons possess numerous distinctive morphological, physiological, and synaptic characteristics, as well as intrinsic and extrinsic connections that distinguish them from other CA regions (see Table 1), and more DEGs that are unique for CA2 regions have been described [11]. Despite the identification of several molecular markers in this area by current studies, our understanding of its functional properties, including its unique physiology, signaling and resilience, and behavioral role, particularly in synaptic plasticity and PD, remains limited.
3. Dopamine and Social Behavior in Parkinson’s Disease
PD is an irreversible neurodegenerative disease [1] characterized by the loss of dopaminergic neurons in the pars compacta of the substantia nigra. Dopamine replacement therapy is the primary treatment for PD, aiming to alleviate symptoms and improve the quality of life [55]. However, the role of dopamine extends beyond motor function. It also plays a crucial role in memory formation, particularly in the hippocampus. Research has shown that dopamine activity in the hippocampus is associated with cognitive function, such as social recognition memory in mice [56] and stimulus generalization in humans [57]. Furthermore, dopamine is critical for social abilities, as evidenced by studies demonstrating that dopamine receptor antagonists can reduce social play in rats [58,59]. Interestingly, there is a complex relationship between stress, social cognition, and the dopamine system [60,61]. Research has shown that the dopamine D1 receptor in the lateral nucleus of the cerebellum (LCN) is involved in spatial navigation and the formation of social recognition memory in both mice and humans [62]. In animal models of autism, an intriguing correlation has been observed between reduced dopamine neuron activity and impaired social preference [63]. Additionally, dopamine neurons in the dorsal raphe nucleus (DRN) have been found to be involved in social isolation and can mediate a “loneliness-like” state in mice [64]. Recently, a study proposed that the dopaminergic metabolic pathway was specifically disrupted in the hippocampus and prefrontal cortex (PFC) of chronically socially defeated and stressed mice [65]. Furthermore, dopamine transporter (DAT) proteins were found to be enriched in patients with social anxiety disorder [66]. Interestingly, a recent study discovered specific cells with firing patterns similar to dopamine D1 receptor-like neurons in the medial prefrontal cortex (mPFC) that are necessary and sufficient for social memory in mice [67]. However, the role of dopamine itself in CA2 function, particularly in the context of PD, requires further investigation.
4. Complex Changes in the Social Behavior of People with Parkinson’s Disease
Indeed, motor, autonomic, and cognitive impairments are commonly observed in PD [68]. Studies have reported social cognitive decline, including perception, language, and decision-making [4], as well as temporal-order memory deficits [5], in the early stages of PD. PD patients typically exhibit rigidity, tremor, and bradykinesia as classical motor symptoms. Notably, PD patients not only experience multiple motor symptoms but also encounter cognitive deficits [69] and other neuropsychological symptoms, such as depression and anxiety [70,71]. Cognitive impairment can also affect executive function, which encompasses higher-level cognitive processes such as decision-making and problem-solving. Moreover, autonomic neurological deficits, including insomnia, constipation, and lower urinary tract symptoms (LUTS), can exacerbate social deficits. Typically, individuals with motor dysfunction alone but normal cognition are diagnosed with PD, whereas those with motor dysfunction and dementia are classified as having PD dementia (PDD) or PD with mild cognitive impairment (PD-MCI). Up to 25% of people with PD have MCI and are at increased risk of developing PDD [72]. It is estimated that 10%–80% of people with PD are likely to develop PDD [73]. Research suggests that social cognitive deficits are present in the early stages of PD and can worsen as the disease progresses [4]. Social cognition includes the perception of facial expressions, personality, behavior, and relationships, and it involves inferring and judging information from social interactions [74]. PD patients exhibit social cognitive impairments in several aspects, including decision-making, emotion, recognition, and empathy. The theory of mind (ToM), which includes affective and cognitive aspects, is the most representative theory of social cognition. It refers to the ability of individuals to understand their own and others’ psychological states and to predict and explain the behavior of others [75]. Studies demonstrate that people with PD score lower on ToM assessments compared to healthy individuals. This suggests that both affective [76,77] and cognitive aspects of ToM are impaired and that these impairments worsen as the disease progresses [78]. Visuospatial abilities contribute significantly to ToM [77]. In addition, research suggests that these impairments are separate from other PD symptoms, such as cognitive function, depression, and motor impairments [79]. Poor performance on advanced ToM tasks in PD patients is associated with executive function deficits [80].
Patients with PD have impaired decision-making, as studies have shown that they have difficulty learning from feedback and making optimal decisions, possibly due to dopamine depletion in the basal ganglia [81]. PD patients have difficulty making appropriate judgments in risky situations [81,82,83] and exhibit impaired social communication skills [84]. In addition, moral decision-making in PD patients differs from that of healthy individuals and is based on ToM [85] rather than stress [86]. Patients with PD and mild cognitive impairment show difficulties with problem-solving [87], suggesting a possible link between social cognitive deficits and executive dysfunction [88,89]. PD patients also have difficulty recognizing negative emotions and showing empathy [90,91,92,93,94]. These deficits may be related to dopaminergic depletion in the limbic system and prefrontal cortex [95,96]. However, there are few studies that showed that dopamine replacement therapy failed to abolish ToM deficits, suggesting that the dopaminergic mechanism may not be involved [97,98].
The intricate alterations in social behavior observed in individuals with PD present a challenge in identifying precise molecular mechanisms or neural circuits, including the hippocampal CA2 region, that contribute to these changes. Further research is necessary to establish a correlation between CA2 functionality and specific social behavioral variations observed in PD patients. Nonetheless, a few studies have indicated a potential involvement of CA2 in human social behavior [99,100,101]. Furthermore, there is evidence that some other diseases with social cognitive deficits, such as schizophrenia, showed apparent changes in human CA2. This collective body of research highlights the significant role of the human hippocampal formation in social cognition [102,103,104,105,106], as summarized in Table 2.
5. Exploring the Role of CA2 in Parkinson’s Disease
As the classical view of PD expands beyond its traditional characterization as a purely motor disorder, it becomes increasingly important to delve into the contributions of non-motor brain regions. Among these regions, the hippocampal formation has been widely studied, and findings indicate that the hippocampus may possess compensatory mechanisms that help mitigate cognitive impairments associated with PD [120]. In parallel, explorations of the CA2 region within the hippocampus have illuminated a noteworthy association between the frequency of cortical Lewy bodies and the extent of neuritic degeneration [121].
The presence of α-synuclein in brainstem nuclei serves as a defining feature of PD and is closely linked to motor impairment. However, it is worth noting that α-synuclein also accumulates in the cerebral cortex and hippocampus, where it plays a crucial role in the development of cognitive abilities [46,47]. This accumulation has been shown to have detrimental effects on synapses by activating neuronal extrasynaptic NMDA receptors (eNMDARs) [49]. Interestingly, within the hippocampus, α-synuclein is particularly enriched in the CA2 region [47]. Notably, postmortem studies have revealed a strong correlation between α-synuclein accumulation in CA2 and dementia in PD and PDD patients [50,51]. In this context, phosphorylated human S129-α-synuclein, which constitutes the majority of α-synuclein in Lewy bodies, has been found to predominantly accumulate in CA2 compared to other regions of the hippocampus in a transgenic PD-like mouse model [122]. Interestingly, in stage III of PD in humans, α-synuclein expression was significantly elevated only in CA2, whereas other hippocampal areas showed lower levels. The highest levels of α-synuclein in CA2 were observed in stages IV and V of PD [51]. However, similar findings were not observed in A53T mice. Notably, another study reported the accumulation of α-synuclein in the stratum lacunosum-moleculare (SLM) of CA2 in mice, which was associated with innervation from the entorhinal cortex (EC) [52]. Furthermore, some researchers have proposed that this type of pathology in CA2 may be related to cholinergic dysfunction [53,54].
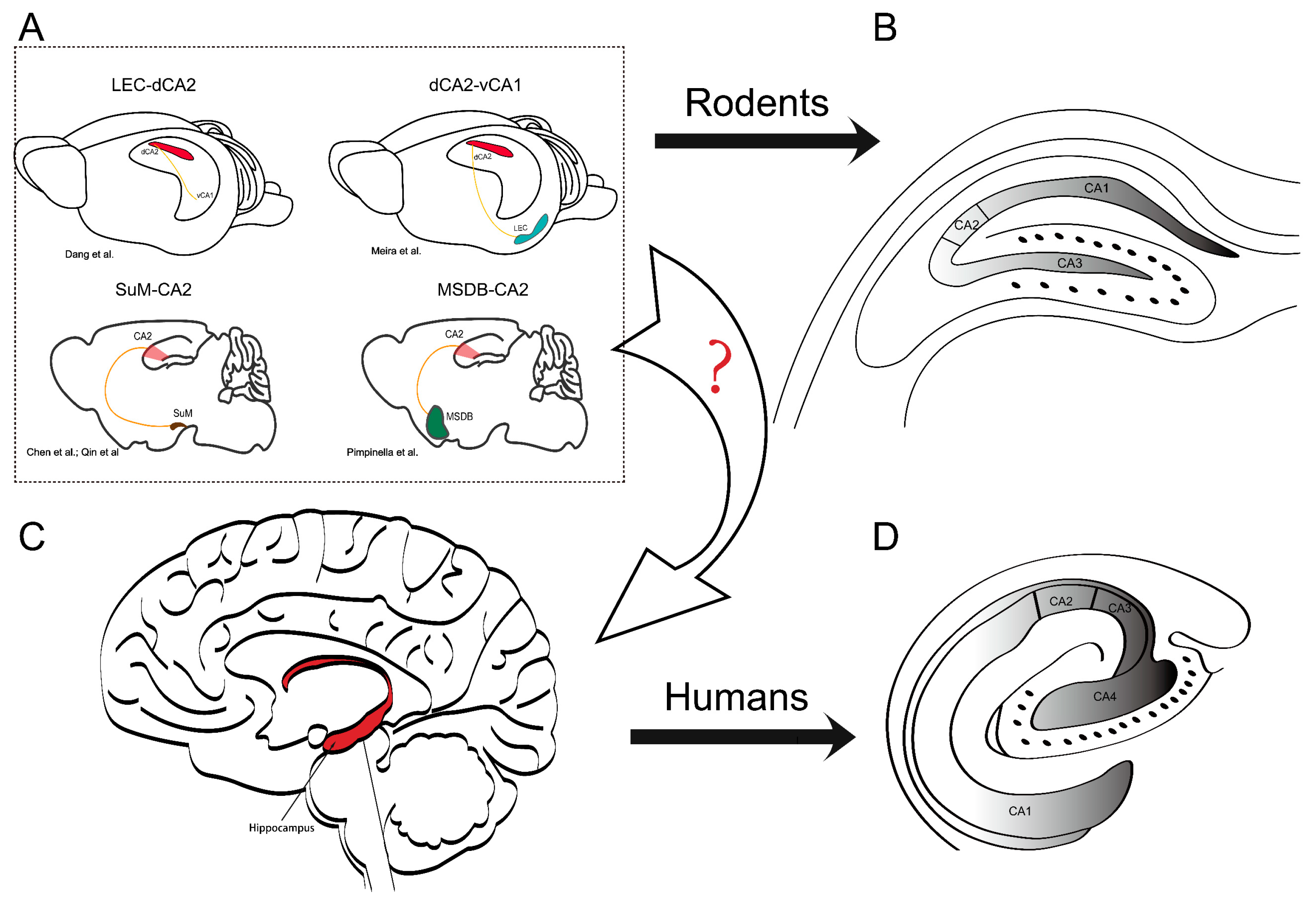
However, a recent study conducted in mice demonstrated that the infusion of α-synuclein fibrils into the CA2–CA3 region did not result in behavioral deficits or cell loss, despite the presence of α-synucleinopathy in this area [45]. However, the model in this study mimics the early stage of PD, suggesting the need to design experiments that address the later stages of PD, when cognitive impairments become more prevalent. On the other hand, it may illustrate the protective role of CA2 in the early stage of this disease. CA2 may not only manifest a unique feature in physiology but also show a different fate in response to different diseases compared to other CA regions. For instance, studies investigating temporal lobe epilepsy in rats [123] and brain injury in humans [124] have consistently shown that pyramidal neurons in CA2 display remarkable resilience to cell death. Additionally, a study focusing on hippocampal sclerosis indicated that neuronal loss in CA2 was less severe compared to other areas of the hippocampus [125]. This suggests that CA2 may possess a distinct resistance to neuronal impairment, possibly due to its robust calcium-handling capabilities, which reduce its vulnerability to cytotoxic events [126]. Furthermore, recent investigations have underscored the importance of considering specific circuits associated with CA2: SuM-CA2 [29,30], lateral entorhinal cortex (LEC)-CA2 [127] and the medial septum-diagonal band of Broca complex (MSDB)-CA2 [128]. However, it remains essential to gain a comprehensive understanding of how these circuits are organized in humans (Figure 1A).
The findings suggest that social cognitive impairments are present in both individuals with PD and animal models of the disease, potentially linked to CA2 function. However, the extent to which CA2 contributes to these cognitive deficits, such as social interaction and temporal ordering, remains uncertain. Furthermore, considering the structural disparities between rodents and humans [129], it is imperative to investigate whether humans exhibit similar functional properties and interbrain connectivity to CA2 as observed in rodents (Figure 1B,C).
The treatment of PD involves various medications, but there is a notable lack of effective options for addressing non-motor symptoms, particularly social disorders. Therefore, it is crucial to identify treatments that can specifically target and alleviate these symptoms without exacerbating motor impairments. In this context, understanding the therapeutic potential of the CA2 region becomes highly relevant. Existing research has examined the effects of specific drugs on the CA2 region (see Table 3). For example, vasopressin, commonly used for diabetes insipidus, has been found to inhibit long-term potentiation (LTP) in the CA2 [130]. Furthermore, dantrolene, a medication for malignant hyperthermia, and ketamine, an anesthetic and antidepressant, have shown promise in rescuing CA2 apoptosis following electroconvulsive seizures (ESC) [131]. Studies in rats have shown that antipsychotics such as haloperidol, clozapine, and olanzapine can reduce the expression of the neuronal glutamate transporter EAAT3 [132] and NMDA receptors in CA2 [133]. In addition, oxytocin, known to improve social cognitive deficits in autism, may exert its effects via somatostatin interneurons in the DG and CA2/CA3 regions [41]. Notably, a human study suggested that long-term treatment with L-DOPA can restore the CA2 volume in patients with PD [134].
In summary, several medications hold potential for improving social memory, enhancing sociability, and reducing social anxiety. Among these is oxytocin, a hormone renowned for its role in promoting social bonding and trust. Studies have suggested that intranasal administration of oxytocin may improve social memory, increase sociability, and reduce social anxiety. Another class of medications, selective serotonin reuptake inhibitors (SSRIs), commonly prescribed as a treatment for depression and anxiety disorders, may also have a positive effect on social anxiety and social functioning. In addition, certain drugs that act as NMDA receptor antagonists, such as ketamine, have shown potential for improving sociability and reducing social anxiety, particularly in individuals with conditions such as autism spectrum disorder [131].
Furthermore, the review of existing literature uncovers the potential impact of specific traditional Chinese medicines (TCMs) on the CA2 region. For example, administration of CS 4-O-sulfation increases the presence of perineuronal networks (PNNs) and excitatory–inhibitory synapses in CA2 [135]. Another chemical compound, dihydroartemisinin, derived from TCM artemisinin, has exhibited protective effects on CA2 neurons against apoptosis induced by lipopolysaccharides (LPSs) [136]. In addition, a TCM compound known as NaoTaiFang has been shown to protect CA2 neurons following cerebral ischemia by increasing the expression of ferroportin (Fpn) and facilitating neuronal iron efflux [137]. Although studies have reported potential benefits of certain TCMs in the context of PD (for comprehensive reviews, refer to [138,139]), it remains unclear whether these TCMs effectively alleviate non-motor symptoms, particularly social disorders. Furthermore, it is necessary to determine whether identified TCMs have an impact on CA2 and the associated social cognitive dysfunction in PD.

{kind=link}
Table 3.
Medications and their effects on the CA2 region.
| Medicine | Indication/Use | Effect on CA2 Region | Refs. |
|---|---|---|---|
| Allopathic Medicines | |||
| Vasopressin | Diabetes insipidus; cardiac arrest | LTP inhibition in EC-CA2 | [130] |
| Dantrolene | MH | Protection of ECS-induced apoptosis in CA2 | [131] |
| Ketamine | Anesthetics | Protect apoptosis induced by ECS in CA2 | [131] |
| Caffeine | N/A | Enhancement of synaptic transmission in CA2 | [39] |
| L-DOPA | PD | Restoration of CA2 volume in PD patients | [134] |
| Fluoxetine | Depression; OCD | Reduction in synaptic protein and GR expression in CA2 | [118] |
| Haloperidol | Schizophrenia; TS | Decrease in EAAT2 and NMDAR in the CA2 | [132,133] |
| Clozapine | Schizophrenia | Decrease in EAAT2 and NMDAR in the CA2 | [132,133] |
| Olanzapine | Schizophrenia | Decrease in NMDAR in the CA2 | [133] |
| Oxytocin | Delivery medication; autism | Rescue of social impairment in an autism model in association with SST neurons in CA2 | [41] |
| Traditional Chinese Medicines (TCMs) | |||
| CS 4-O-sulfation | N/A | Increase in PNNs and excitatory–inhibitory synapses in CA2 | [135] |
| NaoTaiFang | Activating blood and dissolving stasis | Protection of the CA2 neuronal population in cerebral ischemia | [137] |
| Dihydroartemisinin (extract from artemisinin) | Malaria | Protection against LPS-induced apoptosis in CA2 | [136] |
| For more information on specific herbs and TCM formulas for treating PD symptoms, please refer to the following reviews: | [138,139] | ||
LTP: long-term potentiation; ECS: electroconvulsive seizures; CS 4-O-sulfation: chondroitin 4-O-sulfation; PNNs: perineuronal nets; GR: glucocorticoid receptor; EAAT2: excitatory amino acid transporters; NMDAR: N-methyl-D-aspartate receptors; SST: somatostatin interneurons; LPS: lipopolysaccharide; MH: malignant hyperthermia; OCD: obsessive–compulsive disorder; TS: Tourette syndrome.
6. Summary of the Role of CA2 in Non-Motor Symptoms of Parkinson’s Disease
To summarize the putative role of CA2 in the non-motor symptoms of PD, it is important to recognize that although PD is primarily known for its motor symptoms, cognitive impairment and emotional dysregulation are also prevalent. Animal studies have demonstrated that PD-like mice exhibited impaired performance in social behavioral tests, which may indicate some involvement of the CA2 region in social interaction deficits. Furthermore, postmortem studies have revealed CA2 neuronal loss in PD patients, suggesting the vulnerability of this region to neurodegenerative processes. These findings suggest that the CA2 region may play a role in the non-motor symptoms of PD, particularly in social cognition.
Author Contributions
F.Z. performed the literature search, wrote the manuscript, and created the figure. T.B. conceived the structure and content and critically revised the work. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (grant No. 31871076 to T.B., China) and the Shanghai Municipal Science and Technology Major Project (No. 2018SHZDZX01; Shanghai, China), and ZJ Lab, and Shanghai Center for Brain Science and Brain-Inspired Technology.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hague, S.M.; Klaffke, S.; Bandmann, O. Neurodegenerative disorders: Parkinson’s disease and Huntington’s disease. J. Neurol. Neurosurg. Psychiatry 2005, 76, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, K.R.; Healy, D.G.; Schapira, A.H.V. Non-motor symptoms of Parkinson’s disease: Diagnosis and management. Lancet Neurol. 2006, 5, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Aarsland, D. Cognitive impairment in Parkinson’s disease and dementia with Lewy bodies. Parkinsonism Relat. Disord. 2016, 22 (Suppl. S1), S144–S148. [Google Scholar] [CrossRef] [PubMed]
- Palmeri, R.; Buono, V.L.; Corallo, F.; Foti, M.; Di Lorenzo, G.; Bramanti, P.; Marino, S. Nonmotor Symptoms in Parkinson Disease: A Descriptive Review on Social Cognition Ability. J. Geriatr. Psychiatry Neurol. 2017, 30, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.W.; Smith-Petersen, G.A. Time perception and temporal order memory. Acta Psychol. 2014, 148, 173–180. [Google Scholar] [CrossRef]
- van Strien, N.M.; Cappaert, N.L.M.; Witter, M.P. The anatomy of memory: An interactive overview of the parahippocampal–hippocampal network. Nat. Rev. Neurosci. 2009, 10, 272–282. [Google Scholar] [CrossRef]
- Goosens, K.A. Hippocampal regulation of aversive memories. Curr. Opin. Neurobiol. 2011, 21, 460–466. [Google Scholar] [CrossRef]
- Buzsáki, G.; Moser, E.I. Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat. Neurosci. 2013, 16, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Felix-Ortiz, A.C.; Tye, K.M. Amygdala Inputs to the Ventral Hippocampus Bidirectionally Modulate Social Behavior. J. Neurosci. 2014, 34, 586–595. [Google Scholar] [CrossRef] [Green Version]
- Fairént, A.; Regidort, J.; Kruger, L. The Cerebral Cortex of the Mouse (A First Contribution—The “Acoustic” Cortex). Somatosens. Mot. Res. 1992, 9, 3–36. [Google Scholar] [CrossRef]
- Dudek, S.M.; Alexander, G.M.; Farris, S. Rediscovering area CA2: Unique properties and functions. Nat. Rev. Neurosci. 2016, 17, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Lein, E.S.; Callaway, E.M.; Albright, T.D.; Gage, F.H. Redefining the boundaries of the hippocampal CA2 subfield in the mouse using gene expression and 3-dimensional reconstruction. J. Comp. Neurol. 2005, 485, 20426. [Google Scholar] [CrossRef]
- Insausti, R.; Muñoz-López, M.; Insausti, A.M. The CA2 hippocampal subfield in humans: A review. Hippocampus 2023, 33, 712–729. [Google Scholar] [CrossRef]
- Chevaleyre, V.; Siegelbaum, S.A. Strong CA2 Pyramidal Neuron Synapses Define a Powerful Disynaptic Cortico-Hippocampal Loop. Neuron 2010, 66, 560–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohara, K.; Pignatelli, M.; Rivest, A.J.; Jung, H.-Y.; Kitamura, T.; Suh, J.; Frank, D.; Kajikawa, K.; Mise, N.; Obata, Y.; et al. Cell type–specific genetic and optogenetic tools reveal hippocampal CA2 circuits. Nat. Neurosci. 2013, 17, 269–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.E.; Simons, S.B.; Heldt, S.A.; Zhao, M.; Schroeder, J.P.; Vellano, C.P.; Cowan, D.P.; Ramineni, S.; Yates, C.K.; Feng, Y.; et al. RGS14 is a natural suppressor of both synaptic plasticity in CA2 neurons and hippocampal-based learning and memory. Proc. Natl. Acad. Sci. USA 2010, 107, 16994–16998. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, Y.; Hosoya, A.; Yahagi, K.; Ferecskó, A.S.; Yaguchi, K.; Sík, A.; Itakura, M.; Takahashi, M.; Hirase, H. Hippocampal CA3 and CA2 have distinct bilateral innervation patterns to CA1 in rodents. Eur J Neurosci. 2012, 35, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.R.; Parra-Bueno, P.; Smirnov, M.S.; Lustberg, D.J.; Dudek, S.M.; Hepler, J.R.; Yasuda, R. RGS14 Restricts Plasticity in Hippocampal CA2 by Limiting Postsynaptic Calcium Signaling. eNeuro 2018, 5, ENEURO.0353-17. [Google Scholar] [CrossRef] [Green Version]
- Piskorowski, R.A.; Nasrallah, K.; Diamantopoulou, A.; Mukai, J.; Hassan, S.I.; Siegelbaum, S.A.; Gogos, J.A.; Chevaleyre, V. Age-Dependent Specific Changes in Area CA2 of the Hippocampus and Social Memory Deficit in a Mouse Model of the 22q11.2 Deletion Syndrome. Neuron 2016, 89, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Leroy, F.; Park, J.; Asok, A.; Brann, D.H.; Meira, T.; Boyle, L.M.; Buss, E.W.; Kandel, E.R.; Siegelbaum, S.A. A circuit from hippocampal CA2 to lateral septum disinhibits social aggression. Nature 2018, 564, 213–218. [Google Scholar] [CrossRef]
- Young, W.; Li, J.; Wersinger, S.; Palkovits, M. The vasopressin 1b receptor is prominent in the hippocampal area CA2 where it is unaffected by restraint stress or adrenalectomy. Neuroscience 2006, 143, 1031–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitti, F.L.; Siegelbaum, S.A. The hippocampal CA2 region is essential for social memory. Nature 2014, 508, 88–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeVito, L.M.; Konigsberg, R.; Lykken, C.; Sauvage, M.; Young, W.S.; Eichenbaum, H. Vasopressin 1b Receptor Knock-Out Impairs Memory for Temporal Order. J. Neurosci. 2009, 29, 2676–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.S.; Avram, S.K.W.; Cymerblit-Sabba, A.; Song, J.; Young, W.S. Targeted activation of the hippocampal CA2 area strongly enhances social memory. Mol. Psychiatry 2016, 21, 1137–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-T.; Hsieh, T.-Y.; Tsai, T.-C.; Chen, C.-C.; Huang, C.-C.; Hsu, K.-S. Conditional Deletion of Hippocampal CA2/CA3a Oxytocin Receptors Impairs the Persistence of Long-Term Social Recognition Memory in Mice. J. Neurosci. 2017, 38, 1218–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagani, J.H.; Zhao, M.; Cui, Z.; Avram, S.K.W.; Caruana, D.A.; Dudek, S.M.; Young, W.S. Role of the vasopressin 1b receptor in rodent aggressive behavior and synaptic plasticity in hippocampal area CA2. Mol. Psychiatry 2014, 20, 490–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Gerfen, C.R.; Young, W.S., 3rd. Hypothalamic and other connections with dorsal CA2 area of the mouse hippocampus. J. Comp. Neurol. 2013, 521, 1844–1866. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Hernández, V. Synaptic innervation to rat hippocampus by vasopressin-immuno-positive fibres from the hypothalamic supraoptic and paraventricular nuclei. Neuroscience 2013, 228, 139–162. [Google Scholar] [CrossRef]
- Chen, S.; He, L.; Huang, A.J.Y.; Boehringer, R.; Robert, V.; Wintzer, M.E.; Polygalov, D.; Weitemier, A.Z.; Tao, Y.; Gu, M.; et al. A hypothalamic novelty signal modulates hippocampal memory. Nature 2020, 586, 270–274. [Google Scholar] [CrossRef]
- Qin, H.; Fu, L.; Jian, T.; Jin, W.; Liang, M.; Li, J.; Chen, Q.; Yang, X.; Du, H.; Liao, X.; et al. REM sleep-active hypothalamic neurons may contribute to hippocampal social-memory consolidation. Neuron 2022, 110, 4000–4014.e6. [Google Scholar] [CrossRef]
- Dasgupta, A.; Baby, N.; Krishna, K.; Hakim, M.; Wong, Y.P.; Behnisch, T.; Soong, T.W.; Sajikumar, S. Substance P induces plasticity and synaptic tagging/capture in rat hippocampal area CA2. Proc. Natl. Acad. Sci. USA 2017, 114, E8741–E8749. [Google Scholar] [CrossRef] [PubMed]
- Berger, B.; Esclapez, M.; Alvarez, C.; Meyer, G.; Catala, M. Human and monkey fetal brain development of the supramammillary-hippocampal projections: A system involved in the regulation of theta activity. J. Comp. Neurol. 2000, 429, 515–529. [Google Scholar] [CrossRef]
- Meira, T.; Leroy, F.; Buss, E.W.; Oliva, A.; Park, J.; Siegelbaum, S.A. A hippocampal circuit linking dorsal CA2 to ventral CA1 critical for social memory dynamics. Nat. Commun. 2018, 9, 4163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, F.; Brann, D.H.; Meira, T.; Siegelbaum, S.A. Input-Timing-Dependent Plasticity in the Hippocampal CA2 Region and Its Potential Role in Social Memory. Neuron 2017, 95, 1089–1102.e5. [Google Scholar] [CrossRef]
- Alexander, G.M.; Brown, L.Y.; Farris, S.; Lustberg, D.; Pantazis, C.; Gloss, B.; Plummer, N.W.; Jensen, P.; Dudek, S.M. CA2 neuronal activity controls hippocampal low gamma and ripple oscillations. eLife. 2018, 7, e38052. [Google Scholar] [CrossRef]
- Oliva, A.; Fernández-Ruiz, A.; Leroy, F.; Siegelbaum, S.A. Hippocampal CA2 sharp-wave ripples reactivate and promote social memory. Nature 2020, 587, 264–269. [Google Scholar] [CrossRef]
- Srinivas, K.V.; Buss, E.W.; Sun, Q.; Santoro, B.; Takahashi, H.; Nicholson, D.A.; Siegelbaum, S.A. The Dendrites of CA2 and CA1 Pyramidal Neurons Differentially Regulate Information Flow in the Cortico-Hippocampal Circuit. J Neurosci. 2017, 37, 3276–3293. [Google Scholar] [CrossRef]
- McCann, K.E.; Lustberg, D.J.; Shaughnessy, E.K.; Carstens, K.E.; Farris, S.; Alexander, G.M.; Radzicki, D.; Zhao, M.; Dudek, S.M. Novel role for mineralocorticoid receptors in control of a neuronal phenotype. Mol. Psychiatry 2019, 26, 350–364. [Google Scholar] [CrossRef]
- Simons, S.B.; Caruana, D.A.; Zhao, M.; Dudek, S.M. Caffeine-induced synaptic potentiation in hippocampal CA2 neurons. Nat. Neurosci. 2011, 15, 23–25. [Google Scholar] [CrossRef]
- Caruana, D.; Dudek, S.M. Adenosine A1 Receptor-Mediated Synaptic Depression in the Developing Hippocampal Area CA2. Front. Synaptic Neurosci. 2020, 12, 21. [Google Scholar] [CrossRef]
- Bertoni, A.; Schaller, F.; Tyzio, R.; Gaillard, S.; Santini, F.; Xolin, M.; Diabira, D.; Vaidyanathan, R.; Matarazzo, V.; Medina, I.; et al. Oxytocin administration in neonates shapes hippocampal circuitry and restores social behavior in a mouse model of autism. Mol. Psychiatry 2021, 26, 7582–7595. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Lim, Y.J.; Kumar, K.; Baby, N.; Pang, K.L.K.; Benoy, A.; Behnisch, T.; Sajikumar, S. Group III metabotropic glutamate receptors gate long-term potentiation and synaptic tagging/capture in rat hippocampal area CA2. eLife 2020, 9, 55344. [Google Scholar] [CrossRef]
- Robert, V.; Therreau, L.; Davatolhagh, M.F.; Bernardo-Garcia, F.J.; Clements, K.N.; Chevaleyre, V.; Piskorowski, R.A. The mechanisms shaping CA2 pyramidal neuron action potential bursting induced by muscarinic acetylcholine receptor activation. J. Gen. Physiol. 2020, 152, 12462. [Google Scholar] [CrossRef] [PubMed]
- Benoy, A.; Bin Ibrahim, M.Z.; Behnisch, T.; Sajikumar, S. Metaplastic Reinforcement of Long-Term Potentiation in Hippocampal Area CA2 by Cholinergic Receptor Activation. J. Neurosci. 2021, 41, 9082–9098. [Google Scholar] [CrossRef]
- Nouraei, N.; Mason, D.M.; Miner, K.M.; Carcella, M.A.; Bhatia, T.N.; Dumm, B.K.; Soni, D.; Johnson, D.A.; Luk, K.C.; Leak, R.K. Critical appraisal of pathology transmission in the α-synuclein fibril model of Lewy body disorders. Exp. Neurol. 2017, 299, 172–196. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.J.; Grossman, M.; Weintraub, D.; I Hurtig, H.; Duda, J.E.; Xie, S.X.; Lee, E.B.; Van Deerlin, V.M.; Lopez, O.L.; Kofler, J.K.; et al. Neuropathological and genetic correlates of survival and dementia onset in synucleinopathies: A retrospective analysis. Lancet Neurol. 2017, 16, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Adamowicz, D.H.; Roy, S.; Salmon, D.P.; Galasko, D.R.; Hansen, L.A.; Masliah, E.; Gage, F.H. Hippocampal α-Synuclein in Dementia with Lewy Bodies Contributes to Memory Impairment and Is Consistent with Spread of Pathology. J. Neurosci. 2016, 37, 1675–1684. [Google Scholar] [CrossRef] [Green Version]
- Churchyard, A.; Lees, A.J. The relationship between dementia and direct involvement of the hippocampus and amygdala in Parkinson’s disease. Neurology 1997, 49, 1570–1576. [Google Scholar] [CrossRef]
- Trudler, D.; Sanz-Blasco, S.; Eisele, Y.S.; Ghatak, S.; Bodhinathan, K.; Akhtar, M.W.; Lynch, W.P.; Piña-Crespo, J.C.; Talantova, M.; Kelly, J.W.; et al. α-Synuclein Oligomers Induce Glutamate Release from Astrocytes and Excessive Extrasynaptic NMDAR Activity in Neurons, Thus Contributing to Synapse Loss. J. Neurosci. 2021, 41, 2264–2273. [Google Scholar] [CrossRef]
- Kalaitzakis, M.; Pearce, R.; Gentleman, S. Clinical correlates of pathology in the claustrum in Parkinson’s disease and dementia with Lewy bodies. Neurosci. Lett. 2009, 461, 12–15. [Google Scholar] [CrossRef]
- Flores-Cuadrado, A.; Ubeda-Bañon, I.; Saiz-Sanchez, D.; de la Rosa-Prieto, C.; Martinez-Marcos, A. Hippocampal α-synuclein and interneurons in Parkinson’s disease: Data from human and mouse models. Mov. Disord. 2016, 31, 979–988. [Google Scholar] [CrossRef]
- Maki, R.A.; Holzer, M.; Motamedchaboki, K.; Malle, E.; Masliah, E.; Marsche, G.; Reynolds, W.F. Human myeloperoxidase (hMPO) is expressed in neurons in the substantia nigra in Parkinson’s disease and in the hMPO-α-synuclein-A53T mouse model, correlating with increased nitration and aggregation of α-synuclein and exacerbation of motor impairment. Free. Radic. Biol. Med. 2019, 141, 115–140. [Google Scholar] [CrossRef] [PubMed]
- Hall, H.; Reyes, S.; Landeck, N.; Bye, C.; Leanza, G.; Double, K.; Thompson, L.; Halliday, G.; Kirik, D. Hippocampal Lewy pathology and cholinergic dysfunction are associated with dementia in Parkinson’s disease. Brain 2014, 137, 2493–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.K.L.; Chau, T.W.; Lim, E.J.; Ahmed, I.; Chang, R.C.-C.; Kalaitzakis, M.E.; Graeber, M.B.; Gentleman, S.M.; Pearce, R.K.B. Hippocampal CA2 Lewy pathology is associated with cholinergic degeneration in Parkinson’s disease with cognitive decline. Acta Neuropathol. Commun. 2019, 7, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lees, A.J.; Hardy, J.; Revesz, T. Parkinson’s disease. Lancet 2009, 373, 2055–2066. [Google Scholar] [CrossRef] [PubMed]
- Zinn, C.G.; Clairis, N.; Cavalcante, L.E.S.; Furini, C.R.G.; Myskiw, J.D.C.; Izquierdo, I. Major neurotransmitter systems in dorsal hippocampus and basolateral amygdala control social recognition memory. Proc. Natl. Acad. Sci. USA 2016, 113, 1609883113. [Google Scholar] [CrossRef]
- Kahnt, T.; Tobler, P.N. Dopamine regulates stimulus generalization in the human hippocampus. eLife 2016, 5, e12678. [Google Scholar] [CrossRef] [Green Version]
- Trezza, V.; Vanderschuren, L.J.M.J. Divergent Effects of Anandamide Transporter Inhibitors with Different Target Selectivity on Social Play Behavior in Adolescent Rats. Experiment 2008, 328, 343–350. [Google Scholar] [CrossRef]
- Achterberg, M.; Van Kerkhof, L.W.M.; Servadio, M.; van Swieten, M.; Houwing, D.J.; Aalderink, M.; Driel, N.V.; Trezza, V.; Vanderschuren, L.J.M.J. Contrasting Roles of Dopamine and Noradrenaline in the Motivational Properties of Social Play Behavior in Rats. Neuropsychopharmacology 2015, 41, 858–868. [Google Scholar] [CrossRef] [Green Version]
- Barik, J.; Marti, F.; Morel, C.; Fernandez, S.P.; Lanteri, C.; Godeheu, G.; Tassin, J.-P.; Mombereau, C.; Faure, P.; Tronche, F. Chronic Stress Triggers Social Aversion via Glucocorticoid Receptor in Dopaminoceptive Neurons. Science 2013, 339, 332–335. [Google Scholar] [CrossRef]
- Gabriel, P.; Mastracchio, T.-A.; Bordner, K.; Jeffrey, R. Impact of enriched environment during adolescence on adult social behavior, hippocampal synaptic density and dopamine D2 receptor expression in rats. Physiol. Behav. 2020, 226, 113133. [Google Scholar] [CrossRef] [PubMed]
- Locke, T.M.; Soden, M.E.; Miller, S.M.; Hunker, A.; Knakal, C.; Licholai, J.A.; Dhillon, K.S.; Keene, C.D.; Zweifel, L.S.; Carlson, E.S. Dopamine D1 Receptor–Positive Neurons in the Lateral Nucleus of the Cerebellum Contribute to Cognitive Behavior. Biol. Psychiatry 2018, 84, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bariselli, S.; Tzanoulinou, S.; Glangetas, C.; Prévost-Solié, C.; Pucci, L.; Viguié, J.; Bezzi, P.; O’Connor, E.C.; Georges, F.; Lüscher, C.; et al. SHANK3 controls maturation of social reward circuits in the VTA. Nat. Neurosci. 2016, 19, 926–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, G.A.; Nieh, E.H.; Vander Weele, C.M.; Halbert, S.A.; Pradhan, R.V.; Yosafat, A.S.; Glober, G.F.; Izadmehr, E.M.; Thomas, R.E.; Lacy, G.D.; et al. Dorsal Raphe Dopamine Neurons Represent the Experience of Social Isolation. Cell 2016, 164, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; He, Y.; Chen, X.; Tian, Y.; Cheng, K.; Zhang, L.; Wang, Y.; Yang, D.; Wang, H.; Wu, Z.; et al. Validation of the targeted metabolomic pathway in the hippocampus and comparative analysis with the prefrontal cortex of social defeat model mice. J. Neurochem. 2018, 149, 799–810. [Google Scholar] [CrossRef]
- Hjorth, O.R.; Frick, A.; Gingnell, M.; Hoppe, J.M.; Faria, V.; Hultberg, S.; Alaie, I.; Månsson, K.N.T.; Wahlstedt, K.; Jonasson, M.; et al. Expression and co-expression of serotonin and dopamine transporters in social anxiety disorder: A multitracer positron emission tomography study. Mol. Psychiatry 2021, 26, 3970–3979. [Google Scholar] [CrossRef]
- Xing, B.; Mack, N.R.; Guo, K.-M.; Zhang, Y.-X.; Ramirez, B.; Yang, S.-S.; Lin, L.; Wang, D.V.; Li, Y.-C.; Gao, W.-J. A Subpopulation of Prefrontal Cortical Neurons Is Required for Social Memory. Biol. Psychiatry 2020, 89, 521–531. [Google Scholar] [CrossRef]
- Poewe, W. Non-motor symptoms in Parkinson’s disease. Eur. J. Neurol. 2008, 15 (Suppl. S1), 14–20. [Google Scholar] [CrossRef]
- Irwin, D.J.; Lee, V.M.-Y.; Trojanowski, J.Q. Parkinson’s disease dementia: Convergence of α-synuclein, tau and amyloid-β pathologies. Nat. Rev. Neurosci. 2013, 14, 626–636. [Google Scholar] [CrossRef]
- Kehagia, A.A.; Barker, R.A.; Robbins, T.W. Neuropsychological and clinical heterogeneity of cognitive impairment and dementia in patients with Parkinson’s disease. Lancet Neurol. 2010, 9, 1200–1213. [Google Scholar] [CrossRef]
- Wen, M.; Chan, L.L.; Tan, L.C.S.; Tan, E.K. Depression, anxiety, and apathy in Parkinson’s disease: Insights from neuroimaging studies. Eur. J. Neurol. 2016, 23, 1001–1019. [Google Scholar] [CrossRef] [Green Version]
- Aarsland, D.; Bronnick, K.; Williams-Gray, C.; Weintraub, D.; Marder, K.; Kulisevsky, J.; Burn, D.; Barone, P.; Pagonabarraga, J.; Allcock, L.; et al. Mild cognitive impairment in Parkinson disease: A multicenter pooled analysis. Neurology 2010, 75, 1062–1069. [Google Scholar] [CrossRef]
- Hely, M.A.; Reid, W.G.J.; Adena, M.A.; Halliday, G.M.; Morris, J.G.L. The Sydney multicenter study of Parkinson’s disease: The inevitability of dementia at 20 years. Mov. Disord. 2008, 23, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Channon, S. Frontal lobe dysfunction and everyday problem-solving: Social and non-social contributions. Acta Psychol. 2004, 115, 235–254. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.G.; Millett, A.C. The closing of the theory of mind: A critique of perspective-taking. Psychon. Bull. Rev. 2019, 26, 1787–1802. [Google Scholar] [CrossRef] [PubMed]
- Tsuruya, N.; Kobayakawa, M.; Kawamura, M. Is “reading mind in the eyes” impaired in Parkinson’s disease? Park. Relat. Disord. 2011, 17, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Mckinlay, A.; Albicini, M.; Kavanagh, P.S. The effect of cognitive status and visuospatial performance on affective theory of mind in Parkinson’s disease. Neuropsychiatr. Dis. Treat. 2013, 9, 1071–1076. [Google Scholar] [CrossRef] [Green Version]
- Poletti, M.; Vergallo, A.; Ulivi, M.; Sonnoli, A.; Bonuccelli, U. Affective theory of mind in patients with Parkinson’s disease. Psychiatry Clin. Neurosci. 2013, 67, 273–276. [Google Scholar] [CrossRef]
- Bodden, M.E.; Mollenhauer, B.; Trenkwalder, C.; Cabanel, N.; Eggert, K.M.; Unger, M.M.; Oertel, W.H.; Kessler, J.; Dodel, R.; Kalbe, E. Affective and cognitive theory of mind in patients with parkinson’s disease. Park. Relat. Disord. 2010, 16, 466–470. [Google Scholar] [CrossRef]
- Yu, R.-L.; Wu, R.-M.; Chiu, M.-J.; Tai, C.-H.; Lin, C.-H.; Hua, M.-S. Advanced Theory of Mind in patients at early stage of Parkinson’s disease. Park. Relat. Disord. 2012, 18, 21–24. [Google Scholar] [CrossRef]
- Xi, C.; Zhu, Y.; Mu, Y.; Chen, B.; Dong, B.; Cheng, H.; Hu, P.; Zhu, C.; Wang, K. Theory of mind and decision-making processes are impaired in Parkinson’s disease. Behav. Brain Res. 2015, 279, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Mimura, M.; Oeda, R.; Kawamura, M. Impaired decision-making in Parkinson’s disease. Parkinsonism Relat. Disord. 2006, 12, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Kobayakawa, M.; Koyama, S.; Mimura, M.; Kawamura, M. Decision making in Parkinson’s disease: Analysis of behavioral and physiological patterns in the Iowa gambling task. Mov. Disord. 2007, 23, 547–552. [Google Scholar] [CrossRef] [PubMed]
- McNamara, P.; Durso, R.; Brown, A.; Lynch, A. Counterfactual cognitive deficit in persons with Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2003, 74, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Rosen, J.B.; Brand, M.; Polzer, C.; Ebersbach, G.; Kalbe, E. Moral decision-making and theory of mind in patients with idiopathic Parkinson’s disease. Neuropsychology 2013, 27, 562–572. [Google Scholar] [CrossRef]
- Starcke, K.; Polzer, C.; Wolf, O.T.; Brand, M. Does stress alter everyday moral decision-making? Psychoneuroendocrinology 2011, 36, 210–219. [Google Scholar] [CrossRef]
- Anderson, R.J.; Simpson, A.C.; Channon, S.; Samuel, M.; Brown, R.G. Social problem solving, social cognition, and mild cognitive impairment in Parkinson’s disease. Behav. Neurosci. 2013, 127, 184–192. [Google Scholar] [CrossRef]
- Santangelo, G.; Vitale, C.; Trojano, L.; Errico, D.; Amboni, M.; Barbarulo, A.M.; Grossi, D.; Barone, P. Neuropsychological correlates of theory of mind in patients with early Parkinson’s disease. Mov. Disord. 2011, 27, 98–105. [Google Scholar] [CrossRef]
- Esteves, S.; Gleichgerrcht, E.; Torralva, T.; Chade, A.; Arévalo, G.G.; Gershanik, O.; Manes, F.; Roca, M. Performance of Patients with Early Parkinson Disease on an Executive and Social Cognition Battery. Cogn. Behav. Neurol. 2018, 31, 142–150. [Google Scholar] [CrossRef]
- De Risi, M.; Di Gennaro, G.; Picardi, A.; Casciato, S.; Grammaldo, L.G.; D’Aniello, A.; Lanni, D.; Meletti, S.; Modugno, N. Facial emotion decoding in patients with Parkinson’s disease. Int. J. Neurosci. 2017, 128, 71–78. [Google Scholar] [CrossRef]
- Multani, N.; Taghdiri, F.; Anor, C.J.; Varriano, B.; Misquitta, K.; Tang-Wai, D.F.; Keren, R.; Fox, S.; Lang, A.E.; Vijverman, A.C.; et al. Association Between Social Cognition Changes and Resting State Functional Connectivity in Frontotemporal Dementia, Alzheimer’s Disease, Parkinson’s Disease, and Healthy Controls. Front. Neurosci. 2019, 13, 1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narme, P.; Bonnet, A.-M.; Dubois, B.; Chaby, L. Understanding facial emotion perception in Parkinson’s disease: The role of configural processing. Neuropsychologia 2011, 49, 3295–3302. [Google Scholar] [CrossRef] [PubMed]
- Pietschnig, J.; Schröder, L.; Ratheiser, I.; Kryspin-Exner, I.; Pflüger, M.; Moser, D.; Auff, E.; Pirker, W.; Pusswald, G.; Lehrner, J. Facial emotion recognition and its relationship to cognition and depressive symptoms in patients with Parkinson’s disease. Int. Psychogeriatrics 2016, 28, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Sedda, A.; Petito, S.; Guarino, M.; Stracciari, A. Identification and intensity of disgust: Distinguishing visual, linguistic and facial expressions processing in Parkinson disease. Behav. Brain Res. 2017, 330, 30–36. [Google Scholar] [CrossRef]
- Lawrence, A.D.; Goerendt, I.K.; Brooks, D.J. Impaired recognition of facial expressions of anger in Parkinson’s disease patients acutely withdrawn from dopamine replacement therapy. Neuropsychologia 2007, 45, 65–74. [Google Scholar] [CrossRef]
- Cohen, H.; Gagné, M.-H.; Hess, U.; Pourcher, E. Emotion and object processing in Parkinson’s disease. Brain Cogn. 2010, 72, 457–463. [Google Scholar] [CrossRef]
- Péron, J.; Vicente, S.; Leray, E.; Drapier, S.; Drapier, D.; Cohen, R.; Biseul, I.; Rouaud, T.; Le Jeune, F.; Sauleau, P.; et al. Are dopaminergic pathways involved in theory of mind? A study in Parkinson’s disease. Neuropsychologia 2009, 47, 406–414. [Google Scholar] [CrossRef]
- Roca, M.; Torralva, T.; Gleichgerrcht, E.; Chade, A.; Arévalo, G.G.; Gershanik, O.; Manes, F. Impairments in Social Cognition in Early Medicated and Unmedicated Parkinson Disease. Cogn. Behav. Neurol. 2010, 23, 152–158. [Google Scholar] [CrossRef]
- Heinrichs, M.; von Dawans, B.; Domes, G. Oxytocin, vasopressin, and human social behavior. Front. Neuroendocr. 2009, 30, 548–557. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Lindenberg, A.; Domes, G.; Kirsch, P.; Heinrichs, M. Oxytocin and vasopressin in the human brain: Social neuropeptides for translational medicine. Nat. Rev. Neurosci. 2011, 12, 524–538. [Google Scholar] [CrossRef]
- Montanez-Miranda, C.; Bramlett, S.N.; Hepler, J.R. RGS14 expression in CA2 hippocampus, amygdala, and basal ganglia: Implications for human brain physiology and disease. Hippocampus 2022, 33, 166–181. [Google Scholar] [CrossRef]
- Quiroga, R.Q. Concept cells: The building blocks of declarative memory functions. Nat. Rev. Neurosci. 2012, 13, 587–597. [Google Scholar] [CrossRef]
- Cao, R.; Lin, C.; Brandmeir, N.J.; Wang, S. A human single-neuron dataset for face perception. Sci. Data 2022, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- Montagrin, A.; Saiote, C.; Schiller, D. The social hippocampus. Hippocampus 2017, 28, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Trinkler, I.; King, J.A.; Doeller, C.F.; Rugg, M.D.; Burgess, N. Neural bases of autobiographical support for episodic recollection of faces. Hippocampus 2009, 19, 718–730. [Google Scholar] [CrossRef] [PubMed]
- Banker, S.M.; Pagliaccio, D.; Ramphal, B.; Thomas, L.; Dranovsky, A.; Margolis, A.E. Altered structure and functional connectivity of the hippocampus are associated with social and mathematical difficulties in nonverbal learning disability. Hippocampus 2020, 31, 79–88. [Google Scholar] [CrossRef]
- Raam, T.; McAvoy, K.M.; Besnard, A.; Veenema, A.H.; Sahay, A. Hippocampal oxytocin receptors are necessary for discrimination of social stimuli. Nat. Commun. 2017, 8, 2001. [Google Scholar] [CrossRef] [Green Version]
- Diethorn, E.J.; Gould, E. Postnatal development of hippocampal CA2 structure and function during the emergence of social recognition of peers. Hippocampus 2022, 33, 208–222. [Google Scholar] [CrossRef]
- Raghuraman, R.; Navakkode, S.; Sajikumar, S. Alteration of hippocampal CA2 plasticity and social memory in adult rats impacted by juvenile stress. Hippocampus 2023, 33, 745–758. [Google Scholar] [CrossRef]
- Maletta, T.; Palummieri, M.; Correa, J.; Holahan, M.R. Preadolescent exposure to a sexually mature, unrelated male rat reduces postadolescent social recognition memory and CA2 c-Fos labeling. Front. Behav. Neurosci. 2023, 17, 1104866. [Google Scholar] [CrossRef]
- Lisgaras, C.P.; Oliva, A.; Mckenzie, S.; LaFrancois, J.; Siegelbaum, S.A.; Scharman, H.E. Hippocampal area CA2 controls seizure dynamics, interictal EEG abnormalities and social comorbidity in mouse models of temporal lobe epilepsy. BioRxiv 2023. [Google Scholar] [CrossRef]
- Rey, C.C.; Robert, V.; Bouisset, G.; Loisy, M.; Lopez, S.; Cattaud, V.; Lejards, C.; Piskorowski, R.A.; Rampon, C.; Chevaleyre, V.; et al. Altered inhibitory function in hippocampal CA2 contributes in social memory deficits in Alzheimer’s mouse model. iScience 2022, 25, 103895. [Google Scholar] [CrossRef] [PubMed]
- Carstens, K.E.; Lustberg, D.J.; Shaughnessy, E.K.; McCann, K.E.; Alexander, G.M.; Dudek, S.M. Perineuronal net degradation rescues CA2 plasticity in a mouse model of Rett syndrome. J. Clin. Investig. 2021, 131, jci137221. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, S.; Rey, C.C.; Therreau, L.; Fanton, A.; Massotte, D.; Verret, L.; Piskorowski, R.A.; Chevaleyre, V. Maturation of PNN and ErbB4 Signaling in Area CA2 during Adolescence Underlies the Emergence of PV Interneuron Plasticity and Social Memory. Cell Rep. 2019, 29, 1099–1112.e4. [Google Scholar] [CrossRef] [Green Version]
- Wersinger, S.R.; Kelliher, K.R.; Zufall, F.; Lolait, S.J.; O’Carroll, A.-M.; Young, W.S. Social motivation is reduced in vasopressin 1b receptor null mice despite normal performance in an olfactory discrimination task. Horm. Behav. 2004, 46, 638–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Rojas, J.; de Solis, C.A.; Leroy, F.; Kandel, E.R.; Siegelbaum, S.A. A direct lateral entorhinal cortex to hippocampal CA2 circuit conveys social information required for social memory. Neuron 2022, 110, 1559–1572.e4. [Google Scholar] [CrossRef]
- Cope, E.C.; Wang, S.H.; Waters, R.C.; Gore, I.R.; Vasquez, B.; Laham, B.J.; Gould, E. Activation of the CA2-ventral CA1 pathway reverses social discrimination dysfunction in Shank3B knockout mice. Nat. Commun. 2023, 14, 1750. [Google Scholar] [CrossRef]
- Gemmel, M.; Hazlett, M.; Bögi, E.; De Lacalle, S.; Hill, L.A.; Kokras, N.; Hammond, G.L.; Dalla, C.; Charlier, T.D.; Pawluski, J.L. Perinatal fluoxetine effects on social play, the HPA system, and hippocampal plasticity in pre-adolescent male and female rats: Interactions with pre-gestational maternal stress. Psychoneuroendocrinology 2017, 84, 159–171. [Google Scholar] [CrossRef]
- Chevaleyre, V.; Piskorowski, R. Hippocampal Area CA2: An Overlooked but Promising Therapeutic Target. Trends Mol. Med. 2016, 22, 645–655. [Google Scholar] [CrossRef]
- Nagano-Saito, A.; Habak, C.; Mejía-Constaín, B.; Degroot, C.; Monetta, L.; Jubault, T.; Bedetti, C.; Lafontaine, A.-L.; Chouinard, S.; Soland, V.; et al. Effect of mild cognitive impairment on the patterns of neural activity in early Parkinson’s disease. Neurobiol. Aging 2014, 35, 223–231. [Google Scholar] [CrossRef]
- Pang, C.C.; Kiecker, C.; O’Brien, J.T.; Noble, W.; Chang, R.C. Ammon’s Horn 2 (CA2) of the Hippocampus: A Long-Known Region with a New Potential Role in Neurodegeneration. Neurosci. 2018, 25, 167–180. [Google Scholar] [CrossRef] [Green Version]
- Cinar, E.; Yalcin-Cakmakli, G.; Saka, E.; Ulusoy, A.; Yuruker, S.; Elibol, B.; Tel, B.C. Modelling cognitive deficits in Parkinson’s disease: Is CA2 a gateway for hippocampal synucleinopathy? Exp. Neurol. 2020, 330, 113357. [Google Scholar] [CrossRef]
- Norwood, B.A.; Bumanglag, A.V.; Osculati, F.; Sbarbati, A.; Marzola, P.; Nicolato, E.; Fabene, P.F.; Sloviter, R.S. Classic hippocampal sclerosis and hippocampal-onset epilepsy produced by a single “cryptic” episode of focal hippocampal excitation in awake rats. J. Comp. Neurol. 2010, 518, 3381–3407. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, W.L.; Dhillon, K.; Harper, L.; Espin, J.; MacIntosh, T.K.; Smith, D.H.; Graham, D.I. There Is Differential Loss of Pyramidal Cells from the Human Hippocampus with Survival after Blunt Head Injury. J. Neuropathol. Exp. Neurol. 2003, 62, 272–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steve, T.A.; Jirsch, J.D.; Gross, D.W. Quantification of subfield pathology in hippocampal sclerosis: A systematic review and meta-analysis. Epilepsy Res. 2014, 108, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Simons, S.B.; Escobedo, Y.; Yasuda, R.; Dudek, S.M. Regional differences in hippocampal calcium handling provide a cellular mechanism for limiting plasticity. Proc. Natl. Acad. Sci. USA 2009, 106, 14080–14084. [Google Scholar] [CrossRef] [PubMed]
- Dang, R.; Zhou, Y.; Zhang, Y.; Liu, D.; Wu, M.; Liu, A.; Jia, Z.; Xie, W. Regulation of Social Memory by Lateral Entorhinal Cortical Projection to Dorsal Hippocampal CA2. Neurosci. Bull. 2022, 38, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Pimpinella, D.; Mastrorilli, V.; Giorgi, C.; Coemans, S.; Lecca, S.; Lalive, A.L.; Ostermann, H.; Fuchs, E.C.; Monyer, H.; Mele, A.; et al. Septal cholinergic input to CA2 hippocampal region controls social novelty discrimination via nicotinic receptor-mediated disinhibition. eLife 2021, 10, 65580. [Google Scholar] [CrossRef]
- Duvernoy, H.; Françoise, C.; Fatterpekar, G.; Naidich, T.H. The Human Hippocampus, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Chafai, M.; Corbani, M.; Guillon, G.; Desarménien, M.G. Vasopressin Inhibits LTP in the CA2 Mouse Hippocampal Area. PLoS ONE 2012, 7, e49708. [Google Scholar] [CrossRef] [Green Version]
- Gursoy, I.D.; Barun, S.; Erdem, R.; Keskin, U.; Kiziltas, M.; Atilla, P.; Muftuoglu, S.; Yuce, D.; Narin, F.; Ertunc, M.; et al. Investigation of the possible protective effects of ketamine and dantrolene on the hippocampal apoptosis and spatial learning in rats exposed to electroconvulsive seizures as a model of status epilepticus. Turk. Neurosurg. 2020, 30, 871–884. [Google Scholar] [CrossRef]
- Schmitt, A.; Zink, M.; Petroianu, G.; May, B.; Braus, D.F.; Henn, F.A. Decreased gene expression of glial and neuronal glutamate transporters after chronic antipsychotic treatment in rat brain. Neurosci. Lett. 2003, 347, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Krzystanek, M.; Bogus, K.; Palasz, A.; Krzystanek, E.; Worthington, J.J.; Wiaderkiewicz, R. Effects of long-term treatment with the neuroleptics haloperidol, clozapine and olanzapine on immunoexpression of NMDA receptor subunits NR1, NR2A and NR2B in the rat hippocampus. Pharmacol. Rep. 2015, 67, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Györfi, O.; Nagy, H.; Bokor, M.; Moustafa, A.A.; Rosenzweig, I.; Kelemen, O.; Kéri, S. Reduced CA2–CA3 Hippocampal Subfield Volume Is Related to Depression and Normalized by l-DOPA in Newly Diagnosed Parkinson’s Disease. Front. Neurol. 2017, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Joffrin, A.M.; Zhao, Y.; Miller, G.M.; Zhang, G.C.; Oka, Y.; Hsieh-Wilson, L.C. Chondroitin 4- O -sulfation regulates hippocampal perineuronal nets and social memory. Acad. Sci. 2023, 120, e2301312120. [Google Scholar] [CrossRef]
- Gao, Y.; Cui, M.; Zhong, S.; Feng, C.; Nwobodo, A.K.; Chen, B.; Song, Y.; Wang, Y. Dihydroartemisinin ameliorates LPS-induced neuroinflammation by inhibiting the PI3K/AKT pathway. Metab. Brain Dis. 2020, 35, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Xia, X.; Wang, G.-Z.; Shi, Y.-M.; Ge, J.-W. Naotaifang extract treatment results in increased ferroportin expression in the hippocampus of rats subjected to cerebral ischemia. Mol. Med. Rep. 2015, 11, 4047–4052. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Zhang, J.; Wang, C.; Chai, Y.-H.; Wu, A.-G.; Huang, N.-Y.; Wang, L. The pathogenesis and treatment mechanism of Parkinson’s disease from the perspective of traditional Chinese medicine. Phytomedicine 2022, 100, 154044. [Google Scholar] [CrossRef]
- Chen, J.; Xu, J.; Huang, P.; Luo, Y.; Shi, Y.; Ma, P. The potential applications of traditional Chinese medicine in Parkinson’s disease: A new opportunity. Biomed. Pharmacother. 2022, 149, 112866. [Google Scholar] [CrossRef]
Figure 1.
Interbrain connectivity to CA2 and differences in the localization and orientation of the hippocampal formation in rodents and humans. (A) The sketch shows the hippocampal formation and the localization of basic brain-wide connectivity associated with afferents to CA2. The figure highlights four brains with distinct connectivity patterns. In addition, recent studies highlight the need to clarify the organization of these specific circuits in humans: lateral entorhinal cortex (LEC)-CA2 [127], dCA2-vCA1 (d: dorsal, v: ventral) [33], supramammillary area (SuM)-CA2 [29,30], and medial septum–diagonal band of the Broca complex (MSDB)-CA2 [128]. These circuits play a critical role in hippocampal function, and understanding their organization (arrow with “?”) in humans may have implications for various neurological and psychiatric disorders. (B) The CA regions in a rodent hippocampal slice. (C) Human brain and location of the hippocampus. The red area indicates the hippocampal formation. (D) The CA regions in the human hippocampal formation appear to be mirrored with respect to the orientation in the rodent. The figure panels (B,D) from the reference [129] were modified to emphasize the different orientation of the hippocampal formation in humans and rodents.
Figure 1.
Interbrain connectivity to CA2 and differences in the localization and orientation of the hippocampal formation in rodents and humans. (A) The sketch shows the hippocampal formation and the localization of basic brain-wide connectivity associated with afferents to CA2. The figure highlights four brains with distinct connectivity patterns. In addition, recent studies highlight the need to clarify the organization of these specific circuits in humans: lateral entorhinal cortex (LEC)-CA2 [127], dCA2-vCA1 (d: dorsal, v: ventral) [33], supramammillary area (SuM)-CA2 [29,30], and medial septum–diagonal band of the Broca complex (MSDB)-CA2 [128]. These circuits play a critical role in hippocampal function, and understanding their organization (arrow with “?”) in humans may have implications for various neurological and psychiatric disorders. (B) The CA regions in a rodent hippocampal slice. (C) Human brain and location of the hippocampus. The red area indicates the hippocampal formation. (D) The CA regions in the human hippocampal formation appear to be mirrored with respect to the orientation in the rodent. The figure panels (B,D) from the reference [129] were modified to emphasize the different orientation of the hippocampal formation in humans and rodents.

Table 1.
Proteins that are highly expressed in CA2 neurons and their functions.
| Name | Function | References |
|---|---|---|
| PCP4 | Identification of the DG and CA2 regions | [12,15] |
| RGS14 | Restriction of CA2 synaptic plasticity | [15,16,18] |
| STEP | LTP inhibition at EC-CA2 synapses | [15] |
| A1R | LTD enhancement at SC-CA2 synapses | [39,40] |
| AVPR1B | Enhancement of synaptic potentiation at SC-CA2 synapses Facilitation of social behavior | [21,24,26] |
| OXTR | Enhancement of synaptic potentiation at SC-CA2 synapses Facilitation of social behavior | [26,41] |
| MRs; | Facilitation of CA2-dependent behaviors | [38] |
| group III mGluRs; | Restriction of CA2 synaptic plasticity | [42] |
| cholinergic receptors | Induction of LTD at SC and EC CA2 synapses | [43,44] |
| Related to PD | ||
| Substance P | Induction of SC and EC-CA2 synaptic plasticity | [31] |
| α-synuclein | Controversial | [45,46,47,48,49,50,51,52,53,54] |
PCP4: Purkinje cell protein 4; RGS14: regulator of G protein signaling 14 protein; STEP: striatum-enriched protein–tyrosine phosphatase; A1R: A1 adenosine receptor; AVPR1B: vasopressin 1b receptor; OXTR: oxytocin receptor; MRs: mineralocorticoid receptors; Group III mGluRs: Group III metabotropic glutamate receptors.
Table 2.
Comparison of rodent and human studies linking molecular targets and hippocampal formation to social behavior.
Table 2.
Comparison of rodent and human studies linking molecular targets and hippocampal formation to social behavior.
| Social Symptoms | Related Molecules or Factors | Species | References |
|---|---|---|---|
| Social Behavior and CA2 | |||
| Social recognition memory | OXT/OXTR | Mouse | [25,41,107] |
| Ageing | Mouse | [108] | |
| AVPR1B | Mouse | [24] | |
| CA2 pyramidal neurons | Mouse | [22] | |
| Juvenile stress | Rat; mouse | [109,110] | |
| High-frequency oscillations in CA2 neurons | Mouse | [111] | |
| PV interneurons and PNN | Mouse | [112,113,114] | |
| Social novelty recognition | SuM-CA2 synapse | Mouse | [29] |
| Social aggression | AVPR1B | Mouse | [20,26] |
| CA2-LS | Mouse | [20] | |
| Social motivation | AVPR1B | Mouse | [115] |
| Social discrimination | LEC-CA2 | Mouse | [116] |
| Shank3B | Mouse | [117] | |
| Sociability and social interaction | Juvenile stress | Rat | [109] |
| Perinatal fluoxetine | Mouse | [118] | |
| Social cognition | Schizophrenia patients; CA2 PV+ interneurons | Human; mouse | [119] |
| Human Hippocampus and Social Behavior | |||
| Social recognition | Healthy people | Human | [104,105] |
| Social function | Connectivity of hippocampus in NVLD patients | Human | [106] |
| Face perception | Healthy people | Human | [103] |
| Social memory: familiar face and name recognition deficits | Case study | Human | [102] |
| Human Social Behavior and CA2-Related Molecular Targets | |||
| Social stress and anxiety; social cognition and social approach; social behavior | Oxytocin; vasopressin | Human | [99,100] |
| Social anxiety; social discrimination; social behavior; social memory | RGS14 | Human | [101] |
OXT: oxytocin; OXTR: oxytocin receptor; AVPR1B: vasopressin 1b receptor; PV: parvalbumin; PNN: perineuronal net; SuM: supramammillary nuclei; LS: lateral septum; LEC: lateral entorhinal cortex; NVLD: nonverbal learning disabilities; RGS14: regulator of G protein signaling 14 protein.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, F.; Behnisch, T. The Enigmatic CA2: Exploring the Understudied Region of the Hippocampus and Its Involvement in Parkinson’s Disease. Biomedicines 2023, 11, 1996. https://doi.org/10.3390/biomedicines11071996
AMA Style
Zhao F, Behnisch T. The Enigmatic CA2: Exploring the Understudied Region of the Hippocampus and Its Involvement in Parkinson’s Disease. Biomedicines. 2023; 11(7):1996. https://doi.org/10.3390/biomedicines11071996
Chicago/Turabian StyleZhao, Fang, and Thomas Behnisch. 2023. "The Enigmatic CA2: Exploring the Understudied Region of the Hippocampus and Its Involvement in Parkinson’s Disease" Biomedicines 11, no. 7: 1996. https://doi.org/10.3390/biomedicines11071996
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.